El azar y la necesidad
Jacques Monod
(1910-1976)
Jacques-Lucien Monod fue un biólogo y bioquímico francés, ganador del Premio
Nobel de Fisiología o Medicina en 1965, compartido con François Jacob y Andre
Lwoff, «por sus descubrimientos referentes al control genético de la síntesis de
enzimas y virus
ÍNDICE
Presentación
Prefacio
Capítulo 1
Extraños objetos
Contenido:
Lo natural y lo artificial
Las dificultades de un programa espacial
Objetos dotados de un proyecto
Máquinas que se construyen a sí mismas
Máquinas que se reproducen
Las propiedades extrañas: invariancia y teleonomía
La «paradoja» de la invariancia
La teleonomía y el principio de objetividad
Capítulo 2
Vitalismos y animismos
Contenido:
La relación de prioridad entre invariancia y teleonomía: dilema fundamental
Vitalismo metafísico
Vitalismo cientista
La «proyección animista» y la «antigua alianza»
El progresismo cientista
La proyección animista en el materialismo dialéctico
Necesidad de una epistemología crítica
Quiebra epistemológica del materialismo dialéctico
La ilusión antropocentrista
La biosfera: acontecimiento singular no deducible de los primeros principios
Capítulo 3
Los demonios de Maxwell
Contenido:
Las proteínas como agentes moleculares de la teleonomía estructural y funcional
Las proteínas-enzimas como catalizadores específicos
Uniones covalentes y no covalentes
El demonio de Maxwell
Capítulo 4
Cibernética microscópica
Contenido:
Coherencia funcional de la maquinaria celular
Proteínas reguladoras y lógica de las regulaciones
Mecanismo de las interacciones alostéricas
Regulación de la síntesis de las enzimas
La noción de gratuidad
«Holismo» y reduccionismo
Capítulo 5
Ontogénesis molecular
Contenido:
La asociación espontánea de las subunidades en las proteínas oligoméricas
Estructuración espontánea de partículas complejas
Morfogénesis microscópica y morfogénesis macroscópica
Formación de las estructuras globulares
La falsa paradoja del «enriquecimiento» epigenético
La ultima ratio de las estructuras teleonómicas
La interpretación del mensaje
Capítulo 6
Invariancia y perturbaciones
Contenido:
Platón y Heráclito
Las invariantes anatómicas
Las invariantes químicas
El ADN como invariante fundamental
La traducción del código
Irreversibilidad de la traducción
Perturbaciones microscópicas
Incertidumbre operacional e incertidumbre esencial
La evolución: creación absoluta y no revelación
Capítulo 7
Evolución
Contenido:
El azar y la necesidad
Riqueza de la fuente de azar
«Paradoja» de la estabilidad de las especies
La irreversibilidad de la evolución y el segundo principio
Origen de los anticuerpos
El comportamiento como orientando las presiones de selección
El lenguaje y la evolución del hombre
La adquisición primaria del lenguaje
La adquisición del lenguaje programado en el desarrollo epigenético del cerebro
Capítulo 8
Las fronteras
Contenido:
Las fronteras actuales del conocimiento biológico
El problema de los orígenes
El enigma del origen del código
La otra frontera: el sistema nervioso central
Funciones del sistema nervioso central
El análisis de las impresiones sensoriales
Innatismo y empirismo
La función de simulación
La ilusión dualista y la presencia del espíritu
Capítulo 9
El reino y las tinieblas
Contenido:
Presiones de selección en la evolución del hombre
Peligros de degradación genética en las sociedades modernas
La selección de la ideas
La exigencia de explicación
Las ontogenias míticas y metafísicas
La ruptura de la «antigua alianza» animista y el mal del alma moderna
Los valores y el conocimiento
La ética del conocimiento
La ética del conocimiento y el ideal socialista
Apéndices
Contenido:
1. Estructura de las proteínas
2. Ácidos nucleico
3. El código genético
4. Sobre la significación del segundo principio de la termodinámica
Notas:
Presentación
Todo lo que existe en el Universo es fruto del azar y de la necesidad
Demócrito
En ese instante sutil en que el hombre vuelve sobre su vida, Sísifo,
regresando a su roca, contempla esa serie de actos inconexos que devienen su
destino, creado por él, unido bajo la mirada de su memoria y en seguida
sellado por su muerte. Así, persuadido del origen completamente humano de todo
lo que es humano, ciego que desea ver y que sabe que la noche no tiene fin,
está siempre en marcha. La roca todavía rueda. ¡Abandono a Sísifo al pie de la
montaña! Siempre toma a encontrar su fardo. Más Sísifo enseña la fidelidad
superior que niega los dioses y conmueve las rocas. El mismo juzga que todo
está bien. Ese universo, en adelante sin dueño, no le parece ni estéril ni
fútil. Cada grano de esa roca, cada destello mineral de esa montaña, plena de
noche, para él forma un mundo. La propia lucha hacia la cumbre basta para
henchir el corazón de un hombre. Hay que imaginar a Sísifo dichoso.
ALBERT CAMUS
Le mythe de Sisyphe
Prefacio
La biología ocupa, entre las ciencias, un lugar a la vez marginal y central.
Marginal en cuanto que el mundo viviente no constituye más que una parte ínfima
y muy «especial» del universo conocido, de suerte que el estudio de los seres
vivos no parece poder lograr jamás la revelación de unas leyes generales,
aplicables fuera de la biosfera. Pero si la ambición última de la ciencia entera
es fundamentalmente, como creo, dilucidar la relación del hombre con el
Universo, entonces es justo reconocer a la biología un lugar central puesto que
es, entre todas las disciplinas, la que intenta ir más directamente al centro de
los problemas que se deben haber resuelto antes de poder proponer el de la
«naturaleza humana», en unos términos que no sean metafísicos.
Así, la biología es, para el hombre, la más significativa de todas las ciencias;
es la que ha contribuido ya, sin duda, más que ninguna otra, a la formación del
pensamiento moderno, profundamente trastornado y definitivamente marcado en
todos los terrenos: filosófico, religioso y político, por el advenimiento de la
teoría de la evolución. Sin embargo, por segura que estuviese desde el fin del
siglo XIX de su validez fenomenológica, la teoría de la evolución, aunque
dominando la biología entera, permanecía como suspendida mientras no se
elaborara una teoría física de
la herencia. La esperanza de conseguirla rápidamente parecía casi quimérica hace
treinta años a pesar de los éxitos de la genética clásica. Sin embargo, es esto
lo que hoy aporta la teoría molecular del código genético. Interpreto aquí la
«teoría del código genético» en un sentido amplio, para incluir no solamente las
nociones relativas a la estructura química del material hereditario y de la
información de la que es portador, sino también los mecanismos moleculares de
expresión, morfogenética y fisiológica de esta información. Definida así, la
teoría del código genético constituye la base fundamental de la biología. Lo que
no significa, desde luego, que las estructuras y funciones complejas de los
organismos puedan ser deducidas de
la teoría, ni siquiera que sean siempre analizables directamente a escala
molecular. (No se puede predecir ni resolver toda la química con la ayuda de la
teoría cuántica, que sin duda constituye, no obstante, la base universal.)
Pero si la teoría molecular del código no puede hoy (y sin duda no podrá jamás)
predecir y resolver toda la biosfera, constituye desde ahora una teoría general
de los sistemas vivientes. No había nada parecido en el conocimiento científico
anterior al advenimiento de la biología molecular. El «secreto de la vida» podía
entonces parecer inaccesible en su mismo principio. Hoy está en gran parte
desvelado. Este considerable acontecimiento parece debería influir enormemente
en el pensamiento contemporáneo desde el momento en que la significación general
y el alcance de la teoría fuesen comprendidos y apreciados más allá del círculo
de los especialistas puros. Espero que el presente ensayo contribuya a ello.
Antes que las nociones mismas de la biología moderna, es su «forma» lo que he
intentado destacar, así como explicar sus relaciones lógicas con otros terrenos
del pensamiento.
Resulta imprudente hoy en día, por parte de un hombre de ciencia, emplear la
palabra «filosofía», aun siendo «natural» en el título (o incluso en el
subtítulo) de una obra. Se tiene la seguridad de que será acogida con
desconfianza por los científicos y, a lo mejor, con condescendencia por los
filósofos. No tengo más que una excusa, pero la creo legítima: el deber que se
impone, hoy más que nunca, a los hombres de ciencia de considerar a su
disciplina dentro del conjunto de la cultura moderna, para enriquecerla no sólo
de conocimientos técnicos importantes, sino también de las ideas salidas de su
ciencia, que puedan considerarse humanamente significativos. La ingenuidad misma
de una visión nueva (la de la ciencia siempre lo es) puede a veces iluminar con
un nuevo día antiguos problemas.
Desde luego hay que evitar toda confusión entre las ideas sugeridas por
la ciencia y la ciencia misma; pero hay que llevar hasta sus límites las
conclusiones que la ciencia autoriza, a fin de revelar su plena significación.
Ejercicio difícil. Yo no pretendo haber realizado la empresa sin caer en
errores. Digamos que la parte estrictamente biológica de este ensayo no es en
absoluto original. No he hecho más que resumir nociones consideradas como
establecidas por la ciencia contemporánea. La importancia relativa atribuida a
diferentes desarrollos, como la elección de los ejemplos propuestos, reflejan,
es cierto, tendencias personales. Incluso capítulos importantes de la biología
no son mencionados. Repito otra vez que este ensayo no pretende exponer toda la
biología sino que intenta extraer la quintaesencia de la teoría molecular del
código. Indudablemente, soy responsable de las generalizaciones ideológicas que
he creído poder deducir. Mas no creo equivocarme al decir que estas
interpretaciones, mientras no salgan del terreno de la epistemología,
encontrarán el asentimiento de la mayoría de los modernos biólogos. Asumo la
total responsabilidad de los desarrollos de orden ético, si no político, que no
he querido eludir, por peligrosos que fuesen o por ingenuos o demasiado
ambiciosos que puedan, a pesar mío, parecer: la modestia conviene al sabio, pero
no a las ideas que posee y que debe defender.
Y aquí tengo la seguridad, tranquilizadora, de hallarme en total acuerdo con
ciertos biólogos contemporáneos cuya obra merece el mayor respeto.
He de solicitar la indulgencia de los biólogos, por ciertos planteamientos que
les parecerán fastidiosos, y la de los no biólogos por la aridez de exposición
de ciertas nociones «técnicas» inevitables. Los apéndices podrán ayudar a
algunos lectores a superar estas dificultades. Más querría insistir sobre el
hecho de que la lectura no es de ningún modo indispensable para quien no tiene
que afrontar directamente las realidades químicas de la biología.
Este ensayo se basa en una serie de conferencias (las «Robbins Lectures»)
pronunciadas en febrero de 1969 en el Pomona College, en California. Agradezco a
la dirección de este instituto por haberme dado la posibilidad de desarrollar,
ante un joven y ardiente público, ciertos temas que, desde largo tiempo, son
para mí objeto de reflexión, mas no de enseñanza. Análogamente, he hecho de
estos temas el motivo de un curso en el Collège de France durante el año escolar
1969-1970. Es una hermosa y preciada institución que autoriza a sus miembros a
rebasar, a veces, los estrictos límites de la enseñanza que se le ha confiado.
Gracias a Guillaume Budé y a François I.
Clos Saint-Jacques
Abril 1970
Capítulo 1
Extraños objetos
Contenido:
Lo natural y lo artificial
Las dificultades de un programa espacial
Objetos dotados de un proyecto
Máquinas que se construyen a sí mismas
Máquinas que se reproducen
Las propiedades extrañas: invariancia y teleonomía
La «paradoja» de la invariancia
La teleonomía y el principio de objetividad
Lo natural y lo artificial
La distinción entre objetos artificiales y objetos naturales nos parece
inmediata y sin ambigüedad. Un peñasco, una montaña, un río o una nube son
objetos naturales; un cuchillo, un pañuelo, un automóvil son objetos
artificiales, artefactos.[1] Cuando
se analicen estos juicios, se verá, sin embargo, que no son inmediatos ni
estrictamente objetivos. Sabemos que el cuchillo ha sido configurado por el
hombre con vistas a una utilización, a una performance[2] considerada
con anterioridad. El objeto materializa la intención preexistente que lo ha
creado y su forma se explica por la performance que
era esperada incluso antes de que se cumpliera. Nada de esto para el río o el
peñasco que sabemos o pensamos han sido configurados por el libre juego de
fuerzas físicas a las que no sabríamos atribuir ningún «proyecto». Todo ello
suponiendo que aceptamos el postulado base del método científico: la Naturaleza
es objetiva y
no proyectiva.
Es, pues, por referencia a nuestra propia actividad, consciente y proyectiva,
por ser nosotros mismos fabricantes de artefactos, que calibramos lo «natural» o
lo «artificial» de un objeto cualquiera. ¿Sería entonces posible definir por
criterios objetivos y generales las características de los objetos artificiales,
productos de una actividad proyectiva consciente, por oposición a los objetos
naturales, resultantes del juego gratuito de las fuerzas físicas? Para
asegurarse de la entera objetividad de los criterios escogidos, lo mejor sería
sin duda preguntarse si, utilizándolos, se podría redactar un programa que
permitiera a una calculadora distinguir un artefacto de un objeto natural.
Un programa así podría encontrar aplicaciones de sumo interés. Supongamos que
una nave espacial deba posarse próximamente en Venus o en Marte; una cuestión
importantísima sería el conocer si estos planetas están o han sido habitados por
seres inteligentes capaces de una actividad proyectiva. Para descubrir tal
actividad, presente o pasada, son evidentemente sus
productos lo
que se deberá reconocer, por diferentes que sean de los frutos de la industria
humana. Desconociéndolo todo de la naturaleza de tales seres, y de los proyectos
que podrían haber concebido, será necesario que el programa no utilice más que
criterios muy generales, basados exclusivamente en la estructura y la forma de
los objetos examinados, prescindiendo de su función eventual.
Vemos que los criterios a emplear serán dos: 1°, regularidad; 2º, repetición.
Mediante el criterio de regularidad se consideraría el hecho de que los objetos
naturales, configurados por el juego de las fuerzas físicas, no presentan casi
nunca estructuras geométricas simples: superficies planas, aristas rectilíneas,
ángulos rectos, simetrías exactas por ejemplo; mientras que los artefactos
presentarían en general tales características, aunque sólo fuera de forma
aproximada y rudimentaria.
El criterio de repetición será sin duda el más decisivo. Materializando un
proyecto, artefactos homólogos, destinados al mismo uso, reproducen
renovadamente, de modo muy aproximado, las intenciones constantes de su creador.
Bajo este punto de vista, el descubrimiento de numerosos objetos de formas
bastante bien definidas sería pues muy significativo.
Tales podrían ser, descritos brevemente, los criterios generales utilizables.
Debe precisarse, además, que los objetos a examinar serían de dimensiones macroscópicas,
pero no microscópicas.
Por «macroscópicas» hemos de entender las dimensiones medibles en centímetros;
por «microscópicas» las dimensiones que se expresan normalmente en Angstrom (1
cm = 108 Angstrom).
Esta precisión es indispensable porque, a escala microscópica, se tendría acceso
a estructuras atómicas o moleculares cuyas geometrías simples y repetitivas no
testimoniarían evidentemente una intención consciente y racional, sino las leyes
químicas.
Las dificultades de un programa espacial
Supongamos el programa escrito y la máquina realizada. Para someter sus performances a
la prueba, no habría nada mejor que hacerla operar sobre objetos terrestres.
Invirtamos nuestras hipótesis, e imaginemos que la máquina ha sido construida
por los expertos de la NASA marciana, deseosos de detectar en la Tierra los
testimonios de una actividad organizada, creadora de artefactos. Y supongamos
que la primera nave marciana aterriza en el bosque de Fontainebleau, por ejemplo
cerca de Barbizon. La máquina examina y compara las dos series de objetos más
destacables de los alrededores: las casas de Barbizon por un lado y las peñas de
Barbizon por otro. Utilizando los criterios de regularidad, de simplicidad
geométrica y de repetición, decidirá fácilmente que las peñas son objetos
naturales, mientras que las casas son artefactos.
Centrando ahora su atención sobre objetos de dimensiones más reducidas, la
máquina examina unos pequeños guijarros, descubriendo al lado de ellos
cristales, por ejemplo de cuarzo. Siguiendo los mismos criterios, deberá
evidentemente llegar a la conclusión de que, si bien los guijarros son
naturales, los cristales de cuarzo son objetos artificiales. Juicio que parece
justificar un «error» en la estructura del programa. «Error» cuyo origen además
es interesante: si los cristales presentan formas geométricas perfectamente
definidas, es que su estructura macroscópica refleja directamente la estructura
microscópica, simple y repetitiva de los átomos o moléculas que los constituyen.
El cristal, en otros términos, es la expresión macroscópica de una estructura
microscópica. Este «error» sería por otra parte fácil de eliminar ya que todas
las estructuras cristalinas posibles son
conocidas.
Pero supongamos que la máquina estudia ahora otro tipo de objeto: una colmena de
abejas silvestres, por ejemplo. Encontraría evidentemente todos los criterios de
un origen artificial: estructuras geométricas simples y repetitivas del panal y
de las células constituyentes, por lo que la colmena sería clasificada en la
misma categoría de objetos que las casas de Barbizon. ¿Qué pensar de este
juicio? Sabemos que la colmena es «artificial» en el sentido que representa el
producto de la actividad de las abejas. Más tenemos buenas razones para creer
que esta actividad es estrictamente automática, actual pero no conscientemente
proyectiva. Además, como buenos naturalistas, consideramos a las abejas como
seres «naturales». ¿No hay pues una contradicción flagrante al considerar como
«artificial» el producto de la actividad automática de un ser «natural»?
Prosiguiendo la encuesta pronto se vería que si hay contradicción no es por
culpa de un error de programación, sino de la ambigüedad de nuestros juicios.
Porque si la máquina examina ahora no la colmena, sino las mismas abejas, no
podrá ver más que objetos artificiales altamente elaborados. El examen más
superficial revelará elementos de simetría simple: bilateral y traslacional.
Además y sobre todo, examinando abeja tras abeja, el programa observará que la
extrema complejidad de su estructura (número y posición de los pelos
abdominales, por ejemplo, o nerviaciones de las alas) se encuentra reproducida
en todos los individuos con una extraordinaria fidelidad. Prueba segura de que
estos seres son los productos de una actividad deliberada, constructiva y del
orden más refinado. La máquina, sobre la base de tan decisivos documentos, no
podría más que señalar a los oficiales de la NASA marciana su descubrimiento, en
la Tierra, de una industria mucho más evolucionada que la suya.
El rodeo que hemos efectuado a través de lo que sólo es en pequeñísima parte
ciencia ficción, estaba destinado a ilustrar la dificultad de definir la
distinción que, sin embargo, nos parece intuitivamente evidente entre objetos
«naturales» y «artificiales». En efecto, sobre la base de criterios
estructurales (macroscópicos) es sin duda imposible llegar a una definición de
lo artificial que, incluyendo todos los «verdaderos» artefactos, como los
productos de la industria humana, excluya objetos tan evidentemente naturales
como las estructuras cristalinas, así como los seres vivientes mismos, que no
obstante querríamos igualmente clasificar entre los sistemas naturales.
Reflexionando sobre la causa de las confusiones (¿aparentes?) a las que conduce
el programa, se pensará sin duda que ellas surgen por la limitación a que hemos
sometido el mismo al ceñirnos a las consideraciones de forma, de estructura, de
geometría, privando de este modo a la noción de objeto artificial de su
contenido esencial: que un objeto de este tipo se define, se explicó al
principio, por la función que está destinado a cumplir, por la performance que
espera su inventor. Sin embargo se verá en seguida que programando en adelante
la máquina para que estudie no sólo la estructura, sino las performances eventuales
de los objetos examinados, se llegará a resultados aún más engañosos.
Objetos dotados de un proyecto
Supongamos por ejemplo que este nuevo programa permite efectivamente a la
máquina analizar correctamente las estructuras y las performances de
dos series de objetos, tales como caballos corriendo en un campo y automóviles
circulando por una carretera. El análisis llevará a la conclusión de que estos
objetos son comparables, en cuanto están concebidos unos y otros para ser
capaces de realizar desplazamientos rápidos, aunque sobre superficies
diferentes, lo que demuestra sus diferencias de estructura. Y si, para tomar
otro ejemplo, proponemos a la máquina comparar las estructuras y las performances del
ojo de un vertebrado con las de un aparato fotográfico, el programa no podrá más
que reconocer las profundas analogías; lentes, diafragma, obturador, pigmentos
fotosensibles: los mismos componentes no pueden haberse dispuesto, en los dos
objetos, más que con vistas a obtener performances muy
parecidas.
He citado este ejemplo, clásico, de adaptación funcional en los seres vivos,
para subrayar lo estéril y arbitrario de querer negar que el órgano natural, el
ojo, representa el término de un «proyecto» (el de captar imágenes) tan claro
como el que llevó a la consecución del aparato fotográfico. Por lo que sería
absurdo no llegar, en un último análisis, a la conclusión de que el proyecto que
«explica» el aparato no sea el mismo que dio al ojo su estructura. Todo
artefacto es un producto de la actividad de un ser vivo que expresa así, y de
forma particularmente evidente, una de las propiedades fundamentales que
caracterizan sin excepción a todos los seres vivos: la de ser objetos
dotados de un proyecto que
a la vez representan en sus estructuras y cumplen con sus performances (tales
como, por ejemplo, la creación de artefactos).
En vez de rehusar esta noción (como ciertos biólogos han intentado hacer), es
por el contrario indispensable reconocerla como esencial a la definición misma
de los seres vivos. Diremos que éstos se distinguen de todas las demás
estructuras de todos los sistemas presentes en el universo por esta propiedad
que llamaremos teleonomía.
Se notará sin embargo que esta condición, aunque necesaria para la definición de
los seres vivos, no es suficiente ya que no propone criterios objetivos que
permitan distinguir los seres vivientes de los artefactos, productos de su
actividad.
No basta con señalar que el proyecto que da vida a un artefacto pertenece al
animal que lo ha creado, y no al objeto artificial. Esta noción evidente es
todavía demasiado subjetiva, y la prueba de ello es que sería difícil de
utilizar en el programa de una calculadora: ¿cómo sabría que el proyecto de
captar imágenes —proyecto representado por un aparato fotográfico— pertenece a
un objeto aparte del aparato mismo? Por el solo examen de la estructura acabada
y el análisis de sus performances,
es posible identificar el proyecto, pero no su autor.
Para lograrlo, es preciso un programa que estudie no sólo el objeto actual, sino
su origen, su historia y, para empezar, su modo de construcción. Nada se opone,
al menos en principio, a que un programa así pueda ser formulado. Aunque incluso
bastante primitivo, este programa permitiría discernir, entre un artefacto por
perfeccionado que fuera y un ser vivo, una diferencia radical. La máquina no
podría en efecto dejar de constatar que la estructura macroscópica de un
artefacto (se trate de un panal, de una presa erigida por castores, de un hacha
paleolítica, o de un vehículo espacial) es el resultado de la aplicación a los
materiales que lo constituyen, de fuerzas exteriores al
mismo objeto. La estructura macroscópica, una vez acabada, no atestigua las
fuerzas de cohesión internas entre átomos o moléculas que constituyen el
material (y no le confieren más que sus propiedades generales de densidad,
dureza, ductilidad, etc.), sino las fuerzas externas que
lo han configurado.
Máquinas que se construyen a sí mismas
El programa, en contrapartida, deberá registrar el hecho de que la estructura de
un ser vivo resulta de un proceso totalmente diferente en cuanto no debe casi
nada a la acción de las fuerzas exteriores, y en cambio lo debe todo, desde la
forma general al menor detalle, a interacciones «morfogenéticas» internas al
mismo objeto. Estructura testimoniando pues un determinismo autónomo, preciso,
riguroso, implicando una «libertad» casi total con respecto a los agentes o a
las condiciones externas, capaces seguramente de trastornar este desarrollo,
pero incapaces de dirigirlo o de imponer al objeto viviente su organización. Por
el carácter autónomo y espontáneo de los procesos morfogenéticos que construyen
la estructura macroscópica de los seres vivos, éstos se distinguen absolutamente
de los artefactos, así como también de la mayoría de los objetos naturales, en
los que la morfología macroscópica resulta en gran parte de la acción de agentes
externos. Esto tiene una excepción: los cristales, cuya geometría característica
refleja las interacciones microscópicas internas al mismo objeto. Por este
criterio tan sólo, los cristales serian pues clasificados junto a los seres
vivientes, mientras que artefactos y objetos naturales, configurados unos y
otros por agentes externos, constituirían otra clase.
Que por este criterio, así como por el de la regularidad y el de la repetición,
sean agrupadas las estructuras cristalinas y las de los seres vivos, podría
hacerse meditar al programador, incluso ignorando la moderna biología: debería
preguntarse si las fuerzas internas que confieren su estructura macroscópica a
los seres vivos no serían de la misma naturaleza que las interacciones
microscópicas responsables de las morfologías cristalinas. Ello es realmente así
y constituye uno de los principales temas desarrollados en los siguientes
capítulos del presente ensayo. Por el momento, buscamos definir por criterios
absolutamente generales las propiedades macroscópicas que diferencian los seres
vivos de todos los demás objetos del universo.
Habiendo «descubierto» que un determinismo interno, autónomo, asegura la
formación de las estructuras extremadamente complejas de los seres vivientes,
nuestro programador, ignorando la biología, pero experto en informática, debería
ver necesariamente que tales estructuras representan una cantidad considerable
de información de la que falta identificar la fuente: porque toda información
expresada, o recibida, supone un emisor.
Máquinas que se reproducen
Admitamos que, prosiguiendo su encuesta, haga en fin su último descubrimiento:
que el emisor de la información expresada en la estructura de un ser vivo es siempre otro
objeto idéntico al primero. Él ha identificado ahora la fuente y descubierto una
tercera propiedad destacable de estos objetos: el poder de reproducir y
transmitir ne
varietur la
información correspondiente a su propia estructura. Información muy rica, ya que
describe una organización excesivamente compleja, pero integralmente conservada
de una generación a la otra. Designaremos esta propiedad con el nombre de reproducción
invariante,
o simplemente invariancia.
Se verá aquí que, por la propiedad de la reproducción invariante, los seres
vivos y las estructuras cristalinas se encuentran una vez más asociadas y
opuestas a los demás objetos conocidos del universo. Se sabe en efecto que
ciertos cuerpos, en solución sobresaturada, no cristalizan, a menos que no se
hayan inoculado a la solución gérmenes de cristales. Además, cuando se trata de
un cuerpo capaz de cristalizar en dos sistemas diferentes, la estructura de los
cristales que aparecerán en la solución será determinada por la de los gérmenes
empleados. Sin embargo, las estructuras cristalinas representan una cantidad de
información muy inferior a la que se transmite de generación en generación en
los seres vivos más simples que conocemos. Este criterio, puramente
cuantitativo, es necesario subrayarlo, permite distinguir a los seres vivientes
de todos los otros objetos, entre los que no se incluyen los cristales.
* * * *
Abandonamos ahora el programador marciano, sumido en sus reflexiones y supuesto
ignorante de la biología. Esta experiencia imaginaria tenía por objeto el
constreñirnos a «redescubrir» las propiedades más generales que caracterizan a
los seres vivos y que los distinguen del resto del universo. Reconocemos ahora
que sabemos la suficiente biología (suponiendo que hoy se la pueda conocer) para
analizar de más cerca e intentar definir de forma más precisa, si es posible
cuantitativa, las propiedades en cuestión. Hemos encontrado tres: teleonomía,
morfogénesis autónoma, invariancia reproductiva.
Las propiedades extrañas: invariancia y teleonomía
De estas tres propiedades, la invariancia reproductiva
es la más fácil de definir cuantitativamente. Ya que se trata de la capacidad de
reproducir una estructura de alto grado de orden, y ya que el grado de orden de
una estructura puede definirse en unidades de información, diremos que el
«contenido de invariancia» de una especie dada es igual a la cantidad de
información que, transmitida de una generación a otra, asegura la conservación
de la norma estructural especifica. Veremos que es posible, mediante ciertas
hipótesis, llegar a una estimación de esta magnitud.
Supuesto que resultará más sencillo comprender más claramente la noción que se
impone con la más inmediata evidencia por el examen de las estructuras y de las performances de
los seres vivos: la de la teleonomía. Noción que, sin embargo, se revela al
análisis profundamente ambigua, ya que implica la idea subjetiva de «proyecto».
Recordemos el ejemplo del aparato fotográfico: si admitimos que la existencia de
este objeto y su estructura realizan el «proyecto» de captar imágenes, debemos
evidentemente admitir que un «proyecto» parecido se cumple en la emergencia del
ojo de un vertebrado.
Mas todo proyecto particular, sea cual sea, no tiene sentido sino como parte de
un proyecto más general. Todas las adaptaciones funcionales de los seres vivos
como también todos los artefactos configurados por ellos cumplen proyectos
particulares que es posible considerar como aspectos o fragmentos de un proyecto
primitivo único, que es la conservación y la multiplicación de la especie.
Para ser más precisos, escogeremos arbitrariamente definir el proyecto
teleonómico esencial como consistente en la transmisión, de una generación a
otra, del contenido de invariancia característico de la especie. Todas las
estructuras, todas las performances,
todas las actividades que contribuyen al éxito del proyecto esencial serán
llamadas «teleonómicas».
Esto permite proponer una definición de principio del «nivel» teleonómico de una
especie. Se puede, en efecto, considerar que todas las estructuras y performances teleonómicas
corresponden a una cierta cantidad de información que debe ser transferida para
que estas estructuras sean realizadas y estas performances cumplidas.
Llamemos a esta cantidad «la información teleonómica». Se puede entonces
considerar que el «nivel teleonómico» de una especie dada corresponde a la
cantidad de información que debe ser transferida, proporcionalmente, por
individuo, para asegurar la transmisión a la generación siguiente del contenido
específico de invariancia reproductiva.
Se verá fácilmente que el cumplimiento de un proyecto teleonómico fundamental
(es decir, la reproducción invariante) pone en marcha, en diferentes especies y
grados de la escala animal, estructuras y performances variadas,
más o menos elaboradas y complejas. Es preciso insistir sobre el hecho de que no
se trata sólo de las actividades directamente ligadas a la reproducción
propiamente dicha, sino de todas las que contribuyen, aunque sea muy
indirectamente, a la sobrevivencia y a la multiplicación de la especie. El
juego, por ejemplo, en los jóvenes de mamíferos superiores, es un elemento
importante de desarrollo físico y de inserción social. Hay pues un valor
teleonómico como participante en la cohesión del grupo, condición de su
supervivencia y de la expansión de la especie. Es el grado de complejidad de
todas estas estructuras o performances,
concebidas para servir al proyecto teleonómico, lo que se trata de averiguar.
Esta magnitud teóricamente definible no es medible en la práctica. Permite al
menos ordenar groseramente diferentes especies o grupos sobre una «escala
teleonómica». Para tomar un ejemplo extremo, imaginemos un poeta enamorado y
tímido que no osa declarar su amor a la mujer que ama y sólo sabe expresar
simbólicamente su deseo en los poemas que le dedica. Supongamos que la dama, al
fin seducida por estos refinados homenajes, consiente en hacer el amor con el
poeta. Sus poemas habrán contribuido al éxito del proyecto esencial y la
información que contenían debe pues ser contabilizada en la suma de las performances teleonómicas
que aseguran la transmisión de la invariancia genética.
Está claro que el éxito del proyecto no comporta ninguna performance análoga
en otras especies animales, en el ratón por ejemplo. Pero, y este punto es
importante, el contenido de invariancia genética es casi el mismo en el ratón y
en el hombre (y en todos los mamíferos). Las
dos magnitudes que hemos intentado definir son pues totalmente distintas.
Esto nos conduce a considerar una cuestión muy importante que concierne a las
relaciones entre las tres propiedades que hemos reconocido como características
de los seres vivos: teleonomía, morfogénesis autónoma e invariancia. El hecho de
que el programa utilizado las haya identificado sucesiva e independientemente no
prueba que no sean simplemente tres manifestaciones de la misma y única
propiedad fundamental y secreta, inaccesible a toda observación directa. Si éste
fuera el caso, distinguir entre estas propiedades, buscar definiciones
diferentes, podría ser ilusorio y arbitrario. Lejos de dar luz sobre los
verdaderos problemas, de centrarse en el «secreto de la vida», de realmente
disecarlo, no estaríamos más que exorcizándolo.
Es absolutamente verdadero que estas tres propiedades están estrechamente
asociadas en todos los seres vivientes. La invariancia genética no se expresa y
no se revela más que a través y gracias a la morfogénesis autónoma de la
estructura que constituye el aparato teleonómico.
Una primera observación se impone: el estatuto de estas tres nociones no es el
mismo. Si la invariancia y la teleonomía son efectivamente «propiedades»
características de los seres vivos, la estructuración espontánea debe más bien
ser considerada como un mecanismo. Veremos además, en los capítulos siguientes,
que este mecanismo interviene tanto en la reproducción de la información
invariante como en la construcción de las estructuras teleonómicas.
Que este mecanismo en definitiva rinda cuenta de las dos propiedades no implica
sin embargo que deban ser confundidas. Es posible, es de hecho metodológicamente
indispensable, distinguirlas y esto por varias razones.
-
Se puede al menos imaginar objetos capaces de reproducción
invariante, incluso desprovistos de todo aparato teleonómico. Las estructuras
cristalinas pueden ser un ejemplo, a un nivel de complejidad muy inferior, por
cierto, al de todos los seres vivos conocidos.
-
La distinción entre teleonomía e invariancia no es una simple abstracción
lógica. Ella está justificada por consideraciones químicas. En efecto, de las
dos clases de macromoléculas biológicas esenciales, una, la de las proteínas,
es responsable de casi todas las estructuras y performances teleonómicas,
mientras que la invariancia genética está ligada exclusivamente a la otra
clase, la de los ácidos nucleicos.
-
Como se verá en el capitulo siguiente, esta distinción es, explícitamente
o no, supuesta en todas las teorías, en todas las construcciones ideológicas
(religiosas, científicas o metafísicas) relativas a la biosfera y a sus
relaciones con el resto del universo.
* * * *
Los seres vivos son objetos extraños. Los hombres, de todos los tiempos, han
debido más o menos confusamente saberlo. El desarrollo de las ciencias de la
naturaleza a partir del siglo XVII, su expansión a partir del siglo XIX, lejos
de borrar esta impresión de extrañeza, la volvían aún más aguda. Respecto a las
leyes físicas que rigen los sistemas macroscópicos, la misma existencia de los
seres vivos parecía constituir una paradoja, violar ciertos principios
fundamentales sobre los que se basa la ciencia moderna. ¿Cuáles exactamente?
Esto no parece aún resuelto. Se trata pues de analizar precisamente la
naturaleza de esa o esas «paradojas». Ello nos dará la ocasión de precisar el
estatuto, respecto a las leyes físicas, de las dos propiedades esenciales que
caracterizan a los seres vivos: la invariancia reproductiva y la teleonomía.
La «paradoja» de la invariancia
La invariancia parece, en efecto, desde el principio, constituir una propiedad
profundamente paradójica, ya que la conservación, la reproducción, la
multiplicación de las estructuras altamente ordenadas parecen incompatibles con
el segundo principio de la termodinámica. Este principio impone, en efecto, que
todo sistema macroscópico no puede evolucionar más que en el sentido de la
degradación del orden que lo caracteriza.[3]
No obstante, esta predicción del segundo principio no es válida, y verificable,
sino considerando la evolución de conjunto de un sistema energéticamente
aislado.
En el seno de un sistema así, en una de sus fases, se podrá observar la
formación y el crecimiento de estructuras ordenadas sin que por tanto la
evolución de conjunto del sistema deje de obedecer al segundo principio. El
mejor ejemplo nos lo da la cristalización de una solución saturada. La
termodinámica de tal sistema es bien conocida. El crecimiento local de orden que
representa el ensamblaje de moléculas inicialmente desordenadas en una red
cristalina perfectamente definida es «pagado» por una transferencia de energía
térmica de la fase cristalina a la solución: la entropía (el desorden) del
sistema en su conjunto aumenta en la cantidad prescrita por el segundo
principio.
Este ejemplo muestra que un crecimiento local de orden, en el seno de un sistema
aislado, es compatible con el segundo principio. Hemos subrayado, sin embargo,
que el grado de orden que representa un organismo, incluso el más simple, es
incomparablemente más elevado que el que define un cristal. Es preciso
preguntarse si la conservación y la multiplicación invariante de tales
estructuras es igualmente compatible con el segundo principio. Es posible
verificarlo por una experiencia en gran modo comparable a la de la
cristalización.
Tomemos un mililitro de agua conteniendo algunos miligramos de azúcar simple,
como la glucosa, así como sales minerales que comprendan los elementos
esenciales partícipes de la composición de los constituyentes químicos de los
seres vivos (nitrógeno, fósforo, azufre, etc.). Sembremos en este medio una
bacteria de la especie Escherichia
coli,
por ejemplo (longitud 2 μ, peso 5 × 10-13 g
aproximadamente). En el espacio de 36 horas la solución contendrá miles de
millones. Constataremos que alrededor del 40% de azúcar ha sido convertido en
constituyentes celulares, mientras que el resto ha sido oxidado a CO2 y
H2O.
Efectuando el experimento en un calorímetro se puede determinar el balance
termodinámico de la operación y constatar que, como en el caso de la
cristalización, la entropía del conjunto del sistema (bacterias + medio) ha
aumentado un poco más que el mínimo prescrito por el segundo principio. Así,
mientras que la estructura extremadamente compleja que representa la célula
bacteriana ha sido no solamente conservada sino multiplicada millares de
millones de veces, la deuda termodinámica que corresponde a la operación ha sido
debidamente regularizada.
No hay, pues, ninguna violación definible o mesurable del segundo principio. Sin
embargo, asistiendo a este fenómeno, nuestra intuición física no puede dejar de
turbarse y de percibir, todavía más que antes del experimento, toda la rareza.
¿Por qué? Porque vemos claramente que este proceso está desviado, orientado en
una dirección exclusiva: la multiplicación de las células. Éstas, ciertamente,
no violan las leyes de la termodinámica, todo lo contrarío. No se contentan con
obedecerlas; las utilizan, como lo haría un buen ingeniero, para cumplir con la
máxima eficacia el proyecto, realizar «el sueño» (F. Jacob) de toda célula:
devenir células.
La teleonomía y el principio de objetividad
Se ensayará, en un próximo capítulo, dar una idea de la complejidad, del
refinamiento y de la eficacia de la maquinaria química necesaria para la
realización de este proyecto que exige la síntesis de varias centenas de
constituyentes orgánicos diferentes, su ensamblaje en varios millares de
especies macromoleculares, la movilización y la utilización, allá donde sea
necesario, del potencial químico liberado por la oxidación del azúcar, la
construcción de los orgánulos celulares. No hay, sin embargo, ninguna paradoja
física en la reproducción invariante de estas estructuras: el precio
termodinámico de la invariancia está pagado, lo más exactamente posible, gracias
a la perfección del aparato teleonómico que, avaro de calorías, alcanza en su
tarea infinitamente compleja un rendimiento raramente igualado por las máquinas
humanas. Este aparato es enteramente lógico, maravillosamente racional,
perfectamente adaptado a su proyecto: conservar y reproducir la norma
estructural. Y ello, no transgrediendo, sino explotando las leyes físicas en
beneficio exclusivo de su idiosincrasia personal. Es la existencia misma de este
proyecto, a la vez cumplido y proseguido por el aparato teleonómico, lo que
constituye el «milagro». ¿Milagro? No, la verdadera cuestión se plantea a otro
nivel, más profundo, que el de las leyes físicas; es de nuestro entendimiento,
de la intuición que tenemos del fenómeno de lo que se trata. No hay en verdad
paradoja o milagro; simplemente una flagrante contradicción epistemológica.
La piedra angular del método científico es el postulado de la objetividad de la
Naturaleza. Es decir, la negativa sistemática de
considerar capaz de conducir a un conocimiento «verdadero» toda interpretación
de los fenómenos dada en términos de causas finales, es decir de «proyecto». Se
puede datar exactamente el descubrimiento de este principio. La formulación, por
Galileo y Descartes, del principio de inercia, no fundaba sólo la mecánica, sino
la epistemología de la ciencia moderna, aboliendo la física y la cosmología de
Aristóteles. Cierto; ni la razón, ni la lógica, ni la experiencia, ni incluso la
idea de su confrontación sistemática habían faltado a los predecesores de
Descartes. Pero la ciencia, tal como la entendemos hoy, no podía constituirse
sobre estas únicas bases. Le faltaba todavía la austera censura planteada por el
postulado de objetividad. Postulado puro, por siempre indemostrable, porque
evidentemente es imposible imaginar una experiencia que pudiera probar la no
existencia de
un proyecto, de un fin perseguido, en cualquier parte de la naturaleza.
Mas el postulado de objetividad es consustancial a la ciencia, ha guiado todo su
prodigioso desarrollo desde hace tres siglos. Es imposible desembarazarse de él,
aunque sólo sea provisionalmente, o en un ámbito limitado, sin salir del de la
misma ciencia.
La objetividad, sin embargo, nos obliga a reconocer el carácter teleonómico de
los seres vivos, a admitir que en sus estructuras y performances realizan
y prosiguen un proyecto. Hay pues allí, al menos en apariencia, una
contradicción epistemológica profunda. El problema central de la biología es
esta contradicción, que se trata de resolver si es que no es más que aparente, o
de declararla radicalmente insoluble si así verdaderamente resulta ser.
Capítulo 2
Vitalismos y animismos
Contenido:
La relación de prioridad entre invariancia y teleonomía: dilema
fundamental
Vitalismo metafísico
Vitalismo cientista
La «proyección animista» y la «antigua alianza»
El progresismo cientista
La proyección animista en el materialismo dialéctico
Necesidad de una epistemología crítica
Quiebra epistemológica del materialismo dialéctico
La ilusión antropocentrista
La biosfera: acontecimiento singular no deducible de los primeros
principios
La relación de prioridad entre invariancia y teleonomía:
dilema fundamental
De la misma manera que las propiedades teleonómicas de los seres vivos parecen
someter a discusión uno de los postulados de base de la teoría moderna del
conocimiento, toda concepción del mundo, filosófica, religiosa, científica,
supone necesariamente una
solución, implícita o no, de este problema. Toda solución, a su vez, sea cual
sea además la motivación, implica de forma igualmente inevitable una hipótesis
en cuanto a la prioridad, causal y temporal, de las dos propiedades
características de los seres vivientes (invariancia y teleonomía), la una
respecto de la otra.
Reservamos para un capítulo ulterior la exposición y las justificaciones de la
hipótesis considerada como única aceptable a los ojos de la ciencia moderna: que
la invariancia precede necesariamente la teleonomía. O, para ser más explícitos,
la idea darwiniana de que la aparición, la evolución, el refinamiento progresivo
de estructuras cada vez más intensamente teleonómicas, es debido a
perturbaciones sobrevenidas a una estructura poseyendo
ya la propiedad de invariancia,
capaz por consecuencia de «conservar el azar» y por ello de someter los efectos
al juego de la selección natural.
Evidentemente, la teoría que aquí esbozo, breve y dogmáticamente, no es la del
mismo Darwin, quien no podía, en su tiempo, tener idea de los mecanismos
químicos de la invariancia reproductiva, ni de la naturaleza de las
perturbaciones que estos mecanismos sufren. Más con ello no se pretende quitar
importancia a la obra de Darwin sino constatar que la teoría selectiva de la
evolución no ha podido tener su completo sentido, toda su precisión y
certidumbre, hasta después, por lo menos, de una veintena de años.
Hasta el presente, la teoría selectiva es la única propuesta que, haciendo de la
teleonomía una propiedad secundaria, derivada de la invariancia considerada como
única primitiva, sea compatible con el postulado de objetividad. Es igualmente
la única en ser no sólo compatible con la física moderna sino en estar
fundamentada en ella, sin restricciones ni adiciones. Es la teoría de la
evolución selectiva la que en definitiva asegura la coherencia epistemológica de
la biología y le da un lugar entre las ciencias de la «Naturaleza objetiva».
Poderoso argumento ciertamente en favor de la teoría, pero que no bastaría para
justificarla.
Todas las demás concepciones que han sido explícitamente propuestas para rendir
cuenta de la rareza de los seres vivientes, o que están implícitamente arropadas
por las ideologías religiosas y la mayoría de los grandes sistemas filosóficos,
suponen la hipótesis inversa: que la invariancia
es protegida, la ontogenia guiada, la evolución orientada por
un principio teleonómico inicial, del que todos esos fenómenos serian
manifestaciones. En el resto de este capítulo analizaré esquemáticamente la
lógica de estas interpretaciones, muy diversas, en apariencia, pero que implican
el abandono, parcial o total, declarado o no, consciente o no, del postulado de
objetividad. Resultará cómodo, para ello, adoptar una clasificación (algo
arbitraria, es cierto) de estas concepciones, según la naturaleza y extensión
supuesta del principio teleonómico que les corresponda.
Se puede así definir por una parte un primer grupo de teorías que admiten un
principio teleonómico que opera exclusivamente en el seno de la biosfera, de la
«materia viviente». Estas teorías, que llamaré vitalistas,
implican pues una distinción radical entre los seres vivos y el universo
inanimado.
Se puede agrupar por otro lado las condiciones que hacen referencia a un
principio teleonómico universal,
responsable de la evolución cósmica y también de la de la biosfera, en cuyo seno
se expresaría solamente de modo más preciso e intenso. Estas teorías ven a los
seres vivos como los productos más elaborados, más perfectos, de una evolución
universalmente orientada que ha desembocado, porque debía hacerlo,
en el hombre y en la humanidad. Estas concepciones que llamaré «animistas» son
bajo muchos puntos de vista más interesantes que las teorías vitalistas a las
que no consagraré más que un breve resumen.[4]
Vitalismo metafísico
Entre las teorías vitalistas, se pueden discernir tendencias muy diversas. Nos
contentaremos aquí con distinguir entre lo que llamaremos el «vitalismo
metafísico» y el «vitalismo cientista».
El más ilustre promotor del vitalismo metafísico ha sido, sin duda, Bergson. Es
sabido que gracias a un estilo seductor, a una dialéctica metafórica desprovista
de lógica más no de poesía, esta filosofía conoció un inmenso éxito. Hoy parece
haber caído en un casi total descrédito, pero antes, en mi juventud, no se podía
esperar tener éxito en el bachillerato sin haber leído La
evolución creadora.
Es preciso anotar que esta filosofía se basa totalmente en cierta idea de la
vida concebida como un «impulso», una «corriente» radicalmente distinta de la
materia inanimada, pero luchando con ella, dificultándola para
obligarla a organizarse. Contrariamente a casi todos los demás vitalismos o
animismos, el de Bergson no es finalista. Rehúsa encerrar la espontaneidad
esencial de la vida en una determinación cualquiera. La evolución, que se
identifica al impulso vital, no puede tener ni causas finales ni causas
eficientes. El hombre es el estado supremo al que la evolución ha llegado, pero
sin haberlo buscado o previsto. Es más bien la manifestación y la prueba de la
total libertad del impulso creador.
A esta concepción está asociada otra, considerada por Bergson como fundamental:
la inteligencia racional es un instrumento de conocimiento especialmente
adaptado para el dominio de la materia inerte, pero totalmente incapaz de
aprehender los fenómenos de la vida. Sólo el instinto, consustancial al impulso
vital, puede dar una intuición directa, global. Todo discurso analítico y
racional sobre la vida está pues desprovisto de sentido, o más bien fuera de
lugar. El alto desarrollo de la inteligencia racional en el Homo
sapiens,
ha llevado consigo un grave y lamentable empobrecimiento de sus poderes de
intuición, de los que hoy hemos de intentar recobrar sus riquezas.
No intentaré discutir (no se presta, además) esta filosofía. Encerrado en la
lógica y pobre en intuiciones globales, me siento incapaz de hacerlo. Por lo
tanto no considero la actitud de Bergson como insignificante, sino todo lo
contrario. La revuelta, consciente o no, contra lo racional, el respeto
concedido al Id a
expendas del Ego son
signos de nuestro tiempo (sin hablar de la espontaneidad creadora). Si Bergson
hubiese empleado un lenguaje menos claro, un estilo más «profundo», aún se
leería.[5]
Vitalismo cientista
Los vitalistas «científicos» han sido numerosos y entre ellos se encuentran
sabios muy considerados. Pero, mientras hace unos cincuenta años los vitalistas
se reclutaban entre los biólogos (de los que el más conocido, Driesch, abandonó
la embriología por la filosofía), los contemporáneos proceden principalmente de
las ciencias físicas, como M. Elsässer y M. Polany. Es comprensible,
ciertamente, que los físicos hayan sido atraídos, más aún que los biólogos, por
la rareza de los seres vivos. Esquemáticamente resumida, la actitud de M.
Elsässer, por ejemplo, es la siguiente:
Sin duda las extrañas propiedades, invariancia y teleonomía, no violan la
física, pero no
son enteramente explicables con
la ayuda de las fuerzas físicas e interacciones químicas reveladas por el
estudio de los sistemas no vivientes. Es pues indispensable admitir que unos
principios que vendrían a añadirse a
los de la física, operan en la materia viva más no en los sistemas no vivientes
donde, por consiguiente, estos principios electivamente vitales no podían ser
descubiertos. Son estos principios (o leyes biotónicas, empleando la
terminología de Elsässer) los que se tratan de elucidar.
El gran Niels Bohr no descartaba, parece, tales hipótesis.
Pero no pretendía aportar la prueba que confirmara que fuesen necesarios. ¿Lo
son? En definitiva, éste es el problema. Es lo que afirman particularmente
Elsässer y Polany. Lo mínimo que se puede decir es que la argumentación de estos
físicos carece singularmente de rigor y firmeza.
Estos argumentos conciernen respectivamente a cada una de las propiedades
extrañas. En lo que concierne a la invariancia, el mecanismo es hoy en día lo
bastante bien conocido para que se pueda afirmar que ningún principio no físico
es necesario para su interpretación (cf. capítulo VI).
Queda la teleonomía, o más exactamente los mecanismos morfogenéticos que
construyen las estructuras teleonómicas. Es completamente cierto que el
desarrollo embrionario es uno de los fenómenos más milagrosos aparentemente de
toda la biología. Es cierto también que estos fenómenos, admirablemente
descritos por los embriólogos, escapan aún, en gran parte (por razones
técnicas), al análisis genético y bioquímico que sólo, evidentemente, podrían
rendir cuenta. La actitud de los vitalistas considerando que las leyes físicas
son o resultarán, en todo caso, insuficientes para explicar la embriogénesis, no
se basa pues en conocimientos precisos, en observaciones acabadas, sino
solamente en nuestra actual ignorancia. Por el contrario, nuestros conocimientos
relativos a los mecanismos cibernéticos moleculares que regulan la actividad y
el crecimiento celulares, han hecho progresos considerables y contribuirán sin
duda en un próximo futuro a la interpretación del desarrollo. Reservamos para el
capítulo IV la discusión de estos mecanismos, lo que nos dará la ocasión de
volver sobre ciertos argumentos vitalistas. El vitalismo necesita, para
sobrevivir, que subsistan en biología, si no verdaderas paradojas, al menos
algunos «misterios». Los avances de estos últimos veinte años en biología
molecular han reducido singularmente el ámbito de estos misterios, dejando sólo
abierto a las especulaciones vitalistas el campo de la subjetividad: el de la
conciencia. No se corre gran riesgo al prever que, en este dominio por el
momento aún «reservado», estas especulaciones resultarán tan estériles como en
todos donde han actuado hasta el presente.
La «proyección animista» y la «antigua alianza»
Referidas a la infancia de la humanidad, anteriores quizás a la aparición del Homo
sapiens,
las concepciones animistas tienen aún profundas y vivaces raíces en el alma del
hombre moderno.
Nuestros antepasados no podían, sin duda, más que percibir confusamente la
extrañeza de su condición. No tenían las razones que hoy tenemos para sentirse
extraños al universo en el que abrían los ojos. ¿Qué veían primeramente?
Animales, plantas; seres en los que de golpe podían adivinar una naturaleza
parecida a la suya. Las plantas crecen, buscan el sol, mueren; los animales
cazan su presa, atacan a sus enemigos, alimentan y defienden a su prole; los
machos se baten por la posesión de una hembra. Plantas, animales, como el mismo
hombre, se explicaban fácilmente: estos seres tienen un proyecto que es el de
vivir y sobrevivir en su descendencia, aunque el precio sea la muerte. El
proyecto explica el ser y el ser no tiene sentido más que por su proyecto.
Pero alrededor de ellos nuestros antepasados veían también otros objetos
sumamente misteriosos: rocas, ríos, montañas, tormentas, lluvias, cuerpos
celestes. Estos objetos, si existían, era preciso que fuese también por un
proyecto, y que tuviesen un alma para alimentarlo. Así se resolvía para estos
hombres la extrañeza del universo: no existen, en realidad, objetos inanimados,
esto sería incomprensible. En el seno del río, en la cima de la montaña, almas
más secretas alimentaban proyectos más vastos y más impenetrables que aquéllos,
transparentes, de los hombres o de los animales. Así nuestros antepasados sabían
ver en las formas y los acontecimientos de la naturaleza la acción de fuerzas
benévolas u hostiles, pero nunca indiferentes, nunca totalmente extrañas.
El paso esencial del animismo (tal como creo definirlo aquí) consiste en una
proyección a la naturaleza inanimada, de la conciencia que posee el hombre del
funcionamiento intensamente teleonómico de su propio sistema nervioso central.
Es, en otros términos, la hipótesis de que los fenómenos naturales pueden y
deben explicarse en definitiva de la misma manera, por las mismas «leyes», que
la actividad humana subjetiva, consciente y proyectiva. El animismo primitivo
formulaba esta hipótesis con toda ingenuidad, franqueza y precisión, poblando
así la naturaleza de graciosos o temibles mitos que, durante siglos, han
alimentado el arte y la poesía.
Sería equivocado sonreír, incluso con la ternura y el respeto que inspira la
niñez. ¿Creemos acaso que la cultura moderna ha renunciado verdaderamente a la
interpretación subjetiva de la naturaleza? El animismo establecía entre la
Naturaleza y el Hombre una profunda alianza fuera de la cual no se extiende más
que una horrible soledad. ¿Hace falta romper esta ligadura porque el postulado
de objetividad se impone? La historia de las ideas desde el siglo XVII atestigua
los esfuerzos prodigados por los más grandes espíritus para evitar la ruptura,
para forjar de nuevo el anillo de «la antigua alianza». Piénsese en las grandes
tentativas como la de Leibniz, o en el enorme y grave monumento levantado por
Hegel. Pero el idealismo está lejos de haber sido el único refugio de un
animismo cósmico. En el mismo núcleo de ciertas ideologías, que dicen y quieren
estar fundadas sobre la ciencia, se encuentra, bajo una forma más o menos
velada, la proyección animista.
El progresismo cientista
La filosofía biológica de Teilhard de Chardin no merecería detenerse en ella, a
no ser por el sorprendente éxito que ha encontrado hasta en los medios
científicos. Éxito que testimonia la angustia, la necesidad de renovar la
alianza. Teilhard la renueva en efecto sin rodeos. Su filosofía, como la de
Bergson, está enteramente fundada sobre un postulado evolucionista inicial.
Pero, contrariamente a Bergson, admite que la fuerza evolutiva opera en el
universo entero, de las partículas elementales a las galaxias; no hay materia
«inerte», y por lo tanto ninguna distinción de esencia entre materia y vida. El
deseo de presentar esta concepción como «científica», lleva a Teilhard a
fundamentarla sobre una definición nueva de la energía. Ésta estaría de algún
modo distribuida según dos vectores, uno seria (supongo) la energía «ordinaria»
mientras que el otro correspondería a la fuerza de ascendencia evolutiva. La
biosfera y el hombre son los productos actuales de esta ascendencia a lo largo
del vector espiritual de la energía. Esta evolución debe continuar hasta que
toda la energía sea concentrada según este vector: es el punto ω.
Aunque la lógica de Teilhard sea incierta y su estilo laborioso, algunos,
incluso no aceptando enteramente su ideología, reconocen una cierta grandeza
poética. Por mi parte estoy sorprendido por la falta de rigor y de austeridad
intelectual de esta filosofía. Veo, sobre todo, una sistemática complacencia en
querer conciliar, transigir a cualquier precio. Quizá, después de todo, Teilhard
no deja de ser miembro de una orden de la que, tres siglos antes, Pascal atacaba
el laxismo teológico.
La idea de reencontrar la antigua alianza animista con la naturaleza, o de
fundar una nueva, gracias a una teoría universal según la cual la evolución de
la biosfera hasta el hombre estaría en la continuidad sin ruptura de la
evolución cósmica, no ha sido, desde luego, descubierta por Teilhard. Es, en
realidad, la idea central del progresismo cientista del siglo XIX. Se la
encuentra en el centro del positivismo de Spencer y del materialismo dialéctico
de Marx y Engels. La fuerza desconocida e incognoscible que,
según Spencer, opera en todo el universo para crear variedad, coherencia,
especialización, orden, juega exactamente el mismo papel, en definitiva, que la
energía «ascendente» de Teilhard: la historia humana prolonga la evolución
biológica, que forma parte de la evolución cósmica. Gracias a este principio
único, el hombre encuentra al fin en el universo su lugar eminente y necesario,
con la certidumbre del progreso al cual está siempre entregado.
La fuerza diferenciante de Spencer (como la energía ascendente de Teilhard)
representa evidentemente la proyección animista. Para dar un sentido a la
naturaleza, para que el hombre no esté separado por un insondable abismo, para
volverla en fin descifrable e inteligente, era preciso darle un proyecto. A
falta de un alma que alimente este proyecto, se inserta entonces en la
naturaleza una «fuerza» evolutiva, ascendente, de lo que resulta de hecho el
abandono del postulado de objetividad.
La proyección animista en el materialismo dialéctico
Entre las ideologías cientistas del siglo XIX, la más poderosa, la que aún en
nuestros días ejerce una profunda influencia, mucho más allá del círculo sin
embargo vasto de sus adeptos, es evidentemente el marxismo. Así es
particularmente revelador el constatar que, queriendo fundar sobre las leyes de
la misma naturaleza el edificio de sus doctrinas sociales, Marx y Engels hayan
recurrido, ellos también, pero mucho más clara y deliberadamente que Spencer, a
la «proyección animista».
Me parece, en efecto, imposible interpretar de otro modo la famosa «inversión»
por la cual Marx sustituye el materialismo dialéctico a la dialéctica idealista
de Hegel.
El postulado de Hegel: que las leyes más generales que gobiernan el universo en
su evolución son de orden dialéctico, tiene valor en el seno de un sistema que
no reconoce como realidad permanente y auténtica más que al espíritu. Si todos
los acontecimientos, todos los fenómenos, no son más que manifestaciones
parciales de una idea que se piensa, es legítimo buscar en la experiencia
subjetiva del movimiento del pensamiento la expresión más inmediata de las leyes
universales, y ya que el pensamiento procede dialécticamente, está claro que las
«leyes de la dialéctica» gobiernan la naturaleza entera. Pero conservar intactas
estas «leyes» subjetivas, para hacer de ellas las de un universo puramente
material, es efectuar la proyección animista con toda claridad, con todas sus
consecuencias, comenzando por el abandono del postulado de objetividad.
Ni Marx, ni Engels han analizado con detalle, para intentar justificarla, la
lógica de esta inversión de la dialéctica. Pero a partir de los numerosos
ejemplos de aplicación que da particularmente Engels (en el Anti-Dühring y
en la Dialéctica
de la Naturaleza),
se puede intentar reconstruir el pensamiento profundo de los fundadores del
materialismo dialéctico. Las articulaciones esenciales serían las siguientes:
-
El modo de existencia de la materia es el movimiento.
-
El universo, definido como la totalidad de la materia, única existente,
está en un estado de perpetua evolución.
-
Todo conocimiento verdadero del universo contribuye a la inteligencia de
esta evolución.
-
Pero este conocimiento no es obtenido más que en la interacción, evolutiva
y causa de evolución, entre el hombre y la materia (o más exactamente el
«resto» de la materia). Todo conocimiento verdadero es pues «práctico».
-
La conciencia corresponde a esta interacción cognitiva. El pensamiento
consciente refleja por consecuencia el movimiento del mismo universo.
-
Ya que, por lo tanto, el pensamiento es parte y reflejo del movimiento
universal, y ya que su movimiento es dialéctico, es preciso que la ley de
evolución del universo sea dialéctica. Lo que explica y justifica el empleo de
términos como contradicción, afirmación, negación, a propósito de fenómenos
naturales.
-
La dialéctica es constructiva (gracias especialmente a la tercera «ley»).
La evolución del universo es pues ascendente y constructiva. Su más alta
expresión es la sociedad humana, la conciencia, el pensamiento, productos
necesarios de esta evolución.
-
Por el nuevo enfoque de la esencia evolutiva de las estructuras del
universo, el materialismo dialéctico aventaja radicalmente al materialismo del
siglo XVIII que, fundado sobre la lógica clásica, no sabía reconocer más que
interacciones mecánicas entre objetos supuestos invariantes y permanecía
incapaz de pensar la evolución.
Se puede seguramente contestar esta reconstitución, negar que corresponde al
pensamiento auténtico de Marx y Engels. Pero esto es, en resumen, secundario. La
influencia de una ideología reside en la significación que permanece en el
espíritu de sus adeptos y la que de ella dan los epígonos. Innumerables textos
prueban que la reconstitución propuesta es legítima, como representando al menos
la «vulgata» del materialismo dialéctico. No citaré más que un texto, muy
significativo en cuanto su autor era un ilustre biólogo moderno, J. B. S.
Haldane. Escribe en su prefacio a la traducción inglesa de la Dialéctica
de la Naturaleza:
«El marxismo considera la ciencia bajo dos aspectos. En primer lugar, los
marxistas estudian la ciencia entre las otras actividades humanas. Muestran cómo
la actividad científica de una sociedad depende de la evolución de sus
necesidades, o sea de sus métodos de producción que la ciencia a su vez
modifica, así como la evolución de sus necesidades. Pero, en segundo lugar, Marx
y Engels no se limitaban a analizar las modificaciones de la sociedad. En la
Dialéctica, descubren las leyes generales del cambio, no sólo en la sociedad y
en el pensamiento humano, sino en el mundo exterior, proyectado
por el pensamiento humano.
Lo que viene a decir que la dialéctica puede ser aplicada a problemas de ciencia
“pura” tanto como a las relaciones sociales de la ciencia.»
El mundo exterior «reflejado por el pensamiento humano»: todo reside en eso, en
efecto. La lógica de la inversión exige evidentemente que este reflejo sea mucho
mayor que una transposición más o menos fiel del mundo exterior. Es
indispensable, para el materialismo dialéctico, que el «Ding an sich», la cosa o
el fenómeno en sí, llegue hasta el nivel de la conciencia sin alteración ni
empobrecimiento, sin que ninguna selección haya operado entre sus propiedades.
Es preciso que el mundo exterior esté literalmente presente en la conciencia de
la integridad total de sus estructuras y de su movimiento.[6]
A esta concepción se podrían oponer sin duda ciertos textos del mismo Marx.
Aunque esto no quite para considerarla indispensable a la coherencia lógica del
materialismo dialéctico, como los epígonos, e incluso los mismos Marx y Engels,
han reconocido. No olvidemos, además, que el materialismo dialéctico es una
adición relativamente tardía al edificio socio-económico ya erigido por Marx.
Adición claramente destinada a hacer del materialismo histórico una «ciencia»
fundamentada sobre las leyes de la naturaleza.
Necesidad de una epistemología crítica
La radical exigencia del «espejo perfecto» explica el encarnizamiento de los
dialécticos materialistas en repudiar cualquier clase de epistemología critica
que será en adelante inmediatamente calificada de «idealista» y de «kantiana».
Se puede seguramente comprender, en cierta medida, esta actitud, de los hombres
del siglo XIX, contemporáneos de la primera gran explosión científica. Puede muy
bien parecer entonces que el hombre, gracias a la ciencia, estuvo dispuesto a
apoderarse directamente de la naturaleza, de apropiarse la substancia misma.
Nadie, por ejemplo, dudaba que la gravitación fuera una ley de la misma
naturaleza, capturada en su intimidad profunda.
Como se sabe, era por un retomo a las fuentes, a las mismas fuentes del
conocimiento, que la segunda etapa de la ciencia, la del siglo XX, debía surgir.
Desde el fin del siglo XIX, la necesidad absoluta de una epistemología crítica
se vuelve evidente como condición misma de la objetividad del conocimiento. No
son en adelante los filósofos los únicos que se libran a esta crítica, sino
también los hombres de ciencia los que son llevados a incorporarla en la misma
trama teórica. Es gracias a esta condición que pueden desarrollarse la teoría de
la relatividad y la mecánica cuántica.
Por otra parte, el progreso de la neurofisiología y de la psicología
experimental comienza a revelarnos al menos algunos de los aspectos del
funcionamiento del sistema nervioso. Lo bastante para que sea evidente que el
sistema nervioso central no puede, y sin duda no debe, expedir a la conciencia
más que una información codificada, transpuesta, encuadrada en unas normas
preestablecidas: es decir asimilada y no simplemente restituida.
Quiebra epistemológica del materialismo dialéctico
La tesis del reflejo puro, del espejo perfecto que no invertiría incluso la
imagen, nos parece pues hoy en día más insostenible que nunca. Pero a decir
verdad no era necesario esperar los adelantos de la ciencia del siglo XX para
ver aparecer las confusiones y contra-sentidos a los que esta ciencia no podía
dejar de conducir. Para iluminar la linterna del pobre Dühring que ya los
denunciaba, el mismo Engels propone numerosos ejemplos de la interpretación
dialéctica de los fenómenos naturales. Citemos el célebre ejemplo del grano de
cebada dado como ilustración de la tercera ley: «Si un grano de cebada encuentra
las condiciones que le son normales, se opera en él una transformación
específica y, bajo la influencia del calor y de la humedad, germina: el grano
desaparece como grano, es negado, reemplazado por la planta nacida de él,
negación del grano. ¿Más cuál es la carrera normal de esta planta? Crece,
florece, se fecunda y produce nuevos granos de cebada, y tan pronto como han
madurado, el tallo se debilita, la planta se niega por su parte. Como resultado
de esta negación de la negación, tenemos de nuevo el grano de cebada del
principio, no único, sino en número de diez, veinte, treinta…» «Ocurre lo mismo,
añade Engels un poco más adelante, en matemáticas: tomemos una magnitud
algebraica cualquiera, por ejemplo a.
Neguémosla, tenemos –a.
Neguemos esta negación multiplicando –a por
–a,
tenemos a2;
es decir la magnitud positiva primitiva, pero de un grado superior…», etc.
Estos ejemplos ilustran sobre todo la amplitud del desastre epistemológico que
resulta de la utilización «científica» de las interpretaciones dialécticas. Los
dialécticos materialistas modernos evitan en general caer en parecidas
tonterías. Pero hacer de la contradicción dialéctica la «ley fundamental» de
todo movimiento, de toda evolución, no deja de ser un intento de sistematizar,
en nombre de la Dialéctica, dos de los más grandes descubrimientos de su tiempo:
el segundo principio de la termodinámica volverla, en fin, descifrable, y
moralmente significante. Es la «proyección animista», siempre reconocible, sean
cuales sean los disfraces.
Interpretación no sólo extraña a la ciencia, sino incompatible con ella, así ha
resultado cada vez que los dialécticos materialistas, saliendo de la pura
verborrea «teórica», han querido esclarecer las vías de la ciencia experimental
con la ayuda de sus concepciones. El mismo Engels (que sin embargo tenía de la
ciencia de su tiempo un profundo conocimiento) había llegado a rechazar, en
nombre de la Dialéctica, dos de los más grandes descubrimientos de su tiempo: el
segundo principio de la termodinámica y (pese a su admiración por Darwin) la
interpretación puramente selectiva de la evolución. Esto en virtud de los mismos
principios por los que Lenin atacaba, y con qué violencia, la epistemología de
Mach; con que Jdanov más tarde ordenaba a los filósofos rusos combatir «a las
diabluras kantianas de la escuela de Copenhague», y por los que Lyssenko acusaba
a los genetistas de sostener una teoría radicalmente incompatible con el
materialismo dialéctico, o sea necesariamente falsa. Pese a las denegaciones de
los genetistas rusos, Lyssenko tenía razón. La teoría del gen como determinante
hereditaria invariante a través de las generaciones, e incluso de las
hibridaciones, es en efecto totalmente inconciliable con los principios
dialécticos. Es por definición una teoría idealista, puesto que reposa sobre un
postulado de invariancia. El hecho de que se conozca hoy en día la estructura
del gen y el mecanismo de su reproducción invariante no cambia nada, porque la
descripción que da la biología moderna es puramente mecanística. Se trata pues
aún, de una concepción relevante del «materialismo vulgar», mecanicista, y por
consecuencia «objetivamente idealista», como lo anotó Althusser en su severo
comentario de mi Lección inaugural en el Collège de France.
* * * *
He pasado revista brevemente, y muy incompletamente, a diversas ideologías o
teorías. Sé puede pensar que doy de ellas una imagen deformada, por ser parcial.
Intentaré justificarme subrayando que en realidad no buscaba más que destacar lo
que estas concepciones admiten, o implican, en lo concerniente a la biología y,
más especialmente, la relación que suponen entre invariancia y teleonomía. Se ha
visto que todas, sin excepción, hacen de un principio teleonómico inicial el
motor de la evolución, sea de la biosfera sola, sea del universo entero. A los
ojos de la teoría científica moderna todas estas concepciones son erróneas, y
esto no solamente por razones de hecho, que serán discutidas concretamente en el
capítulo VI.
La ilusión antropocentrista
En la base de estos errores hay con toda seguridad la ilusión antropocentrista.
La teoría heliocéntrica, la noción de inercia, el principio de objetividad, no
podían bastar para disipar este antiguo espejismo. La teoría de la evolución, en
vez de hacer desaparecer la ilusión, parecía conferirle una nueva realidad
haciendo del hombre no el centro, sino el heredero por siempre esperado,
natural, del universo entero. Dios, en fin, podía morir, reemplazado por este
nuevo y grandioso espejismo. En adelante el último designio de la Ciencia será
formular una teoría unificada que, fundada sobre un corto número de principios,
darán cuenta de la realidad entera, comprendida la biosfera y el hombre. Es en
esta certidumbre exaltante donde se nutre el progresismo cientista del siglo XIX.
Teoría unificada que los dialécticos materialistas creían haber ya formulado.
Porque le parecía atentar a la certidumbre de que el hombre y el pensamiento
humano son los productos necesarios de una ascendencia cósmica, Engels negó
formalmente el segundo principio. Es significativo que lo haga desde la
introducción de la Dialéctica
de la Naturaleza y
que asocie directamente este tema a una predicación cosmológica apasionada por
la que promete si no a la especie humana, al menos al «cerebro pensante», un
eterno retorno. Retorno, en efecto, a uno de los más antiguos mitos de la
humanidad.[7]
La biosfera: acontecimiento singular no deducible de los
primeros principios
Fue preciso esperar a la segunda mitad del siglo XX para que el nuevo espejismo
antropocentrista, incluido en la teoría de la evolución, se desvaneciese. Hoy
podemos, creo, afirmar que una teoría universal, por completos que fueran sus
éxitos en todos sus puntos, no podría nunca contener a la biosfera, a su
estructura, a su evolución como fenómenos deducibles de
los primeros principios.
Esta proposición puede parecer oscura. Busquemos una aclaración. Una teoría
universal debería evidentemente contener, a la vez, a la relatividad, a la
teoría cuántica, a una teoría de las partículas elementales. Siempre que ciertas
condiciones iniciales pudieran ser formuladas, contendría igualmente una
cosmología que prevería la evolución general del Universo. Sabemos sin embargo
que (contrariamente a lo que creía Laplace, y, después de él, la ciencia y la
filosofía «materialista» del siglo XIX) estas previsiones no podrían ser más que
estadísticas. La teoría contendría, sin duda, la clasificación periódica de los
elementos, pero no podría determinar más que la probabilidad de existencia de
cada uno de ellos. Del mismo modo prevería la aparición de objetos tales como
las galaxias o los sistemas planetarios, pero no podría en ningún caso deducir
de sus principios la existencia necesaria de tales objetos, de tal
acontecimiento, de tal fenómeno particular, trátese de la nebulosa de Andrómeda,
del planeta Venus, del monte Everest o de la tormenta de ayer por la tarde. De
una manera general, la teoría prevería la existencia, las propiedades, las
relaciones de ciertas clases de
objetos o de acontecimientos, pero no podría evidentemente prever la existencia,
ni los caracteres distintivos de ningún objeto, de ningún acontecimiento particular.
La tesis que aquí presentaré es que la biosfera no contiene una clase previsible
de objetos o de fenómenos, sino que constituye un acontecimiento particular,
compatible seguramente con los primeros principios, pero no
deducible de
estos principios. Por lo tanto, esencialmente imprevisible.
Espero que se me comprenda bien. Diciendo que los seres vivos, en cuanto clase,
no son previsibles a partir de los primeros principios, no pretendo en ningún
modo sugerir que no son explicables según
estos mismos principios, que en cierto modo transcienden, y que otros
principios, sólo aplicables a ellos, deban ser invocados. La biosfera es, en mi
opinión, imprevisible en el mismo grado que lo es la configuración particular de
los átomos que constituyen este guijarro que tengo en mi mano. Nadie reprocharía
a una teoría universal el no afirmar y prever la existencia de esta
configuración particular de átomos; nos basta que este objeto actual, único y
real, sea compatible con
la teoría. Este objeto no tiene, según la teoría, el deber de existir, mas tiene
el derecho.
Esto nos basta tratándose de un guijarro, pero no si se trata de nosotros
mismos. Nosotros nos queremos necesarios, inevitables, ordenados desde siempre.
Todas las religiones, casi todas las filosofías, una parte de la ciencia,
atestiguan el incansable, heroico esfuerzo de la humanidad negando
desesperadamente su propia contingencia.
Capítulo 3
Los demonios de Maxwell
Contenido:
Las proteínas como agentes moleculares de la teleonomía estructural y
funcional
Las proteínas-enzimas como catalizadores específicos
Uniones covalentes y no covalentes
El demonio de Maxwell
Las proteínas como agentes moleculares de la teleonomía
estructural y funcional
La noción de teleonomía implica la idea de una actividad orientada,
coherente y constructiva.
Por estos criterios, las proteínas deben ser consideradas como los agentes
moleculares esenciales de las performances teleonómicas
de los seres vivos.
-
Los seres vivos son máquinas químicas. El crecimiento y la multiplicación
de todos los organismos exigen que sean cumplidas millares de reacciones
químicas, gracias a las cuales son elaborados los constituyentes esenciales de
las células. Es lo que se llama el «metabolismo». Este metabolismo está
organizado en un gran número de «vías», divergentes, convergentes o cíclicas,
comprendiendo cada cual una secuencia de reacciones. La orientación precisa y
el rendimiento elevado de esta enorme y microscópica actividad química están
asegurados por una cierta clase de proteínas, las enzimas, que representan el
papel de catalizadores específicos.
-
Como una máquina, todo organismo, incluido el más «simple», constituye una
unidad funcional coherente e integrada. Evidentemente, la coherencia funcional
de una máquina química tan compleja, y además autónoma, exige la intervención
de un sistema cibernético que gobierne y controle la actividad química en
numerosos puntos. Estamos lejos aún, sobre todo en los organismos superiores,
de haber elucidado la estructura íntegra de estos sistemas. Hoy se conocen,
sin embargo, muchos elementos, y en todos estos casos se ha comprobado que los
agentes esenciales son proteínas llamadas «reguladoras», que desempeñan, en
suma, el papel de detectores de señales químicas.
-
El organismo es una máquina que se construye a sí misma. Su estructura
macroscópica no le es impuesta por la intervención de fuerzas exteriores. Se
constituye de forma autónoma, gracias a interacciones constructivas internas.
Aunque nuestros conocimientos concernientes a la mecánica del desarrollo sean
totalmente insuficientes, sin embargo se puede, desde ahora, afirmar que las
interacciones constructivas son microscópicas, moleculares, y que las
moléculas en litigio son esencialmente, si no únicamente, proteínas.
Son proteínas, por consecuencia, las que canalizan la actividad de la máquina
química, aseguran la coherencia de su funcionamiento y la construyen. Todas
estas performances teleonómicas
de las proteínas reposan en último lugar sobre las propiedades llamadas
«estéreo-específicas», es decir, su capacidad de «reconocer» a otras moléculas
(comprendidas otras proteínas) según su forma,
que es determinada por su estructura molecular. Se trata, literalmente, de una
propiedad discriminativa (si no «cognitiva») microscópica. Se puede admitir que
toda performance o
estructura teleonómica de un ser vivo, sea cual sea, puede en principio ser
analizada en términos de interacciones estéreo-específicas de una, de varias, o
de numerosas proteínas[8].
Es de la estructura, de la forma de una proteína dada, de la que depende la
discriminación estéreo-específica particular que constituye su función. En la
medida en que se sabría describir el origen y la evolución de esta estructura se
rendiría cuenta también del origen y de la evolución de la performance teleonómica
a la que esté entregada.
En el presente capítulo se discutirá la función catalítica específica de las
proteínas; en el siguiente, la función reguladora, y en el capítulo V, la
función constructora. El problema del origen de las estructuras funcionales será
abordado en este capítulo y vuelto a considerar en el siguiente.
Se puede, en efecto, estudiar las propiedades funcionales de una proteína sin
tener que referirse al detalle de su estructura particular. (No se conoce
todavía hoy, en todos sus detalles, la estructura en el espacio más que de unas
quince proteínas.) No obstante es necesario el repaso de algunos datos
generales.
Las proteínas son moléculas muy grandes, de peso molecular variando entre 10.000
y 1.000.000 o más. Estas macromoléculas están constituidas por la polimerización
secuencial de compuestos de peso molecular cercano a 100, pertenecientes a la
clase de los «aminoácidos». Toda proteína contiene pues de 100 a 10.000
radicales aminoácidos. Sin embargo, estos numerosísimos radicales pertenecen a
sólo 20 especies químicas diferentes[9] que
se encuentran en todos los seres vivientes, desde las bacterias al hombre. Esta
monotonía de composición constituye una de las más patentes ilustraciones del
hecho de que la prodigiosa diversidad de estructuras macroscópicas de
los seres vivos reposa en realidad sobre una profunda y no menos remarcable
unidad de composición y de estructura microscópica.
Volveremos a ello.
Según su forma general, se pueden distinguir dos clases principales de
proteínas:
-
las proteínas llamadas «fibrosas»; son moléculas muy alargadas que juegan
en los seres vivos un papel principalmente mecánico, a la manera del aparejo
de un barco de vela; aunque las propiedades de algunas de ellas (las del
músculo) sean muy interesantes, no las citaremos aquí;
-
las proteínas llamadas «globulares»; son, con mucho, las más numerosas y,
por sus funciones, las más importantes; en estas proteínas las fibras
constituidas por la polimerización secuencial de los aminoácidos están
replegadas sobre sí mismas, de manera extremadamente compleja, confiriendo así
a estas moléculas una estructura compacta, pseudo globular.[10]
Los seres vivos, incluso los más simples, contienen un gran número de proteínas
diferentes. Se puede estimar este número en 2.500 ± 500, para la bacteria Escherichia
coli (5
× 10−13 g
de peso y 2 μ de longitud, aproximadamente). Para los animales superiores, como
por ejemplo el hombre, se barajan cifras que rozan el millón.
Las proteínas-enzimas como catalizadores específicos
Entre los millares de reacciones químicas que contribuyen al desarrollo y a las performances de
un organismo, cada una es provocada electivamente por una proteína-enzima
particular. Se puede, sin simplificar casi, admitir que cada enzima, en el
organismo, ejerce su actividad catalítica en un solo punto del metabolismo. Es
ante todo por su extraordinaria electividad de
acción que las enzimas se distinguen de los catalizadores no biológicos
empleados en laboratorio o en la industria. Entre estos últimos los hay muy
activos, es decir, capaces en muy pequeña cantidad de acelerar considerablemente
diversas reacciones. Ninguno de estos catalizadores, no obstante, se aproxima en
especificidad de acción a la enzima más «vulgar».
Esta especificidad es doble:
-
Cada enzima no cataliza más de un solo tipo de reacción.
-
Entre los cuerpos, a veces muy numerosos en el organismo, susceptibles de
sufrir este tipo de reacción, la enzima, por regla general, no es activa más
que con respecto a uno solo. Algunos ejemplos permitirán aclarar estas
proposiciones.
Existe una enzima (llamada fumarasa) que cataliza la hidratación (adición de
agua) del ácido fumárico en ácido málico:
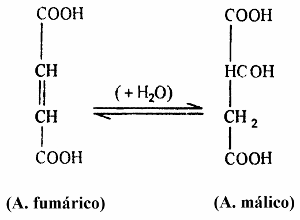
Esta reacción es reversible y la misma enzima cataliza igualmente la
deshidratación del ácido málico en ácido fumárico.
Existe, sin embargo, un isómero geométrico del ácido fumárico, el ácido maleico:
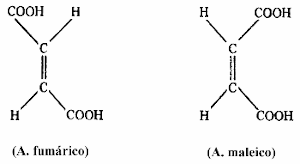
Capaz químicamente de sufrir la misma hidratación. La enzima es totalmente
inactiva en contacto con el segundo.
Pero además existen dos isómeros ópticos del
ácido málico, que poseen un carbono asimétrico:[11]
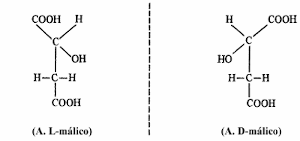
Estos dos cuerpos, imágenes en un espejo el uno del otro, son químicamente
equivalentes y prácticamente inseparables por las técnicas químicas clásicas.
Entre los dos, sin embargo, la enzima ejerce una discriminación absoluta. En
efecto:
-
la enzima deshidrata exclusivamente al ácido L-málico para producir
exclusivamente ácido fumárico;
-
a partir de ácido fumárico, la enzima produce exclusivamente ácido
L-málico y no ácido D-málico.
La discriminación rigurosa ejercida por la enzima entre los isómeros ópticos no
constituye solamente una ilustración espectacular de la especificidad estérica de
las enzimas. En primer lugar se encuentra aquí la explicación del hecho, durante
largo tiempo misterioso, de que entre los numerosos constituyentes químicos
celulares que son disimétricos (de hecho es el caso de la mayoría), uno solo de
los dos isómeros ópticos esté, por regla general, representado en la biosfera.
Pero, en segundo lugar, según el principio muy general de Curie sobre la
conservación de la simetría, el hecho de que a partir de un cuerpo ópticamente
simétrico (ácido fumárico) sea obtenido un cuerpo disimétrico impone que:
-
la enzima constituya la «fuente» de disimetría; o sea, que ella misma sea
ópticamente activa, lo que sucede en este caso;
-
la simetría inicial del substrato se pierda en el curso de su interacción
con la proteína-enzima. Es preciso pues que la reacción de hidratación tenga
lugar en el seno de un «complejo» formado por una asociación temporal entre la
enzima y el substrato; en un complejo así, la simetría inicial del ácido
fumárico sería efectivamente perdida.
La noción de «complejo estéreo-específico», como explicación de la especificidad
así como de la actividad catalítica de las enzimas, es de una importancia
capital. Volveremos sobre ello tras haber discutido otros ejemplos.
Existe (en ciertas bacterias) otra enzima, llamada aspartasa que, también, actúa
exclusivamente sobre el ácido fumárico, a exclusión de todo otro cuerpo,
especialmente de su isómero geométrico, el ácido maleico. La reacción de
«adición sobre el doble enlace» catalizada por esta enzima es casi análoga a la
precedente. Esta vez, no se trata de una molécula de agua, sino de amoniaco, la
que es condensada con el ácido fumárico, para dar un aminoácido, el aspártico:
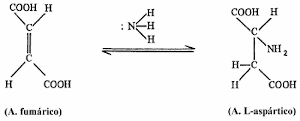
El ácido aspártico posee un carbono asimétrico; es pues ópticamente activo. Como
en el caso precedente, la reacción enzimática produce exclusivamente uno de los
isómeros, el de la serie L, llamado isómero «natural» porque los aminoácidos
entran en la composición de las proteínas perteneciendo todos a la serie L.
Las dos enzimas, aspartasa y fumarasa, discriminan pues estrictamente, no sólo
entre los isómeros ópticos y geométricos de sus substratos y productos, sino
igualmente entre las moléculas de agua y de amoniaco. Se tiende a admitir que
estas últimas moléculas entran, asimismo, en la composición del complejo
estéreo-específico en cuyo seno se produce la reacción de adición y que, en este
complejo, las moléculas están rigurosamente colocadas las unas con respecto a
las otras. Es de esta ordenación de lo que depende el que resulte tanto la
especificidad de acción como la estéreo-especificidad de la reacción.
De los ejemplos precedentes, la existencia de un complejo estéreo-específico
como intermediario de la reacción enzimática no podía ser deducida más que a
título de hipótesis explicativa. En ciertos casos favorables es posible
demostrar directamente la existencia de este complejo. Es el caso de la enzima
llamada β-galactosidasa, que cataliza específicamente la hidrólisis de los
cuerpos que poseen la estructura dada por la fórmula A:
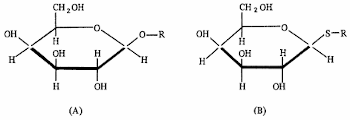
(En estas fórmulas, R representa un radical cualquiera.)
Recordemos que existen numerosos isómeros de tales cuerpos. (16 isómeros
geométricos, que difieren por la orientación relativa de los grupos OH y H en
los carbonos 1 al 5, más los antípodas ópticos de cada uno de estos isómeros.)
La enzima, de hecho, discrimina rigurosamente entre todos estos isómeros, y no
hidroliza más que uno de ellos. Se puede, sin embargo, «engañar» a la enzima
sintetizando «análogos estéticos» de los cuerpos de esta serie, en los cuales el
oxígeno de la unión hidrolizable es reemplazado por azufre (fórmula B). El átomo
de azufre, mayor que el de oxígeno, tiene la misma valencia, y la orientación de
las valencias es la misma en los dos átomos. La forma tridimensional
de estos derivados azufrados es pues prácticamente la misma que la de sus
homólogos de oxígeno. Pero la unión formada por el azufre es mucho más estable
que la del oxigeno. Estos cuerpos no son pues hidrolizados por la enzima. Sin
embargo, se puede demostrar directamente que
forman con la proteína un complejo estéreo-específico.
Tales observaciones no solamente confirman la teoría del complejo, sino
demuestran que una reacción enzimática debe ser considerada como comportando dos
etapas distintas:
-
la formación de un complejo estéreo-específico entre proteína y substrato;
-
la activación catalítica de una reacción en el seno del complejo;
reacción orientada y especificada por la estructura del mismo complejo.
Uniones covalentes y no covalentes
Esta distinción es de una importancia capital y nos va a permitir dar a luz una
de las nociones más importantes de la biología molecular. Pero antes hace falta
recordar que, entre los diferentes tipos de uniones que pueden contribuir a la
estabilidad de un edificio químico, es necesario distinguir dos clases:
-
las uniones llamadas covalentes;
-
las uniones no covalentes.
Las uniones covalentes (a las que se les reserva a menudo el nombre de «unión
química» sensu
stricto)
son debidas a la compartición de orbitales electrónicos entre dos o más átomos.
Las uniones no covalentes son debidas a otros varios tipos de interacciones (que
no implican la compartición de orbitales electrónicos).
No es necesario, para lo que aquí estamos tratando, especificar la naturaleza de
las fuerzas físicas que intervienen en estos diferentes tipos de interacciones.
Subrayemos primeramente que las dos clases de uniones difieren las unas de las
otras por la energía de las asociaciones que aseguran. Simplificando un poco, y
precisando que sólo consideramos aquí reacciones que se producen en fase acuosa,
se puede en efecto admitir que la energía absorbida o liberada, por término
medio, por una reacción que implica uniones covalentes, es del orden de 5 a 20
Kcal (por unión). Para una reacción que implica únicamente uniones no
covalentes, la energía media sería de 1 a 2 Kcal.[12]
Esta importante diferencia pone de manifiesto en parte la diferencia de
estabilidad entre edificios «covalentes» y «no covalentes». Lo esencial, sin
embargo, no está en eso, sino en la diferencia de las energías llamadas de
«activación» puestas en juego en los dos tipos de reacciones. Esta noción es de
una extrema importancia. Para precisarla, recordemos que una reacción que hace
pasar una población molecular de un estado estable dado a otro, debe ser
considerada incluyendo un estado intermediario de energía potencial superior a
la de los estados finales. Se representa a menudo este proceso por una gráfica
cuya abscisa representa la progresión de la reacción y la ordenada la energía
potencial (fig. 1).
Fig. 1. Diagrama de la variación de la energía potencial de las moléculas en
el curso de una reacción. A: estado estable inicial. B: estado estable final. X:
estado intermedio, de energía potencial superior a la de los dos estados
estables. Trazo continuo: reacción covalente. Trazo discontinuo: reacción
covalente en presencia de un catalizador que disminuye la energía de activación.
Trazo punteado: reacción no covalente.
La diferencia de energía potencial entre los estados finales corresponde a la
energía liberada por la reacción. La diferencia entre el estado inicial y el
estado intermedio (llamado «activado») es la energía de activación. Es la
energía que las moléculas deben transitoriamente adquirir
para entrar en reacción. Esta energía, adquirida en una primera etapa, liberada
en la segunda, no figura en el balance termodinámico final. Es de ella, sin
embargo, que depende la velocidad de
la reacción, que será prácticamente nula, a temperatura ordinaria, si la energía
de activación es elevada. Para provocarla, será preciso, pues, o bien aumentar
considerablemente la temperatura (de la que depende la fracción de moléculas que
han adquirido la energía suficiente), o bien emplear un catalizador, cuyo papel
es el de «estabilizar» el estado activado, o sea, reducir la diferencia de
potencial entre este estado y el estado inicial.
Por lo tanto, y éste es el punto importante, en general:
-
la energía de activación de las reacciones covalentes es elevada; su
velocidad es pues muy pequeña o nula a temperatura baja y en ausencia de
catalizadores;
-
la energía de activación de las reacciones no covalentes es muy pequeña,
si no nula; ellas se producen pues espontáneamente, y muy rápidamente a
temperatura baja, y en ausencia de catalizadores.
De ello resulta que las estructuras definidas por interacciones no covalentes no
pueden alcanzar una cierta estabilidad más que poniendo en juego interacciones múltiples.
Además, las interacciones no covalentes no adquieren una energía notable hasta
que los átomos están a distancias muy pequeñas, prácticamente «en contacto» unos
con otros. En consecuencia, dos moléculas (o regiones de moléculas) no podrán
contraer una asociación no covalente a menos que las superficies de las dos
moléculas comprendan áreas
complementarias que
permitan a varios átomos de una entrar en contacto con varios átomos de la otra.
La noción de complejo estéreo-específico no covalente
Si añadimos ahora que los complejos formados por enzima y substrato son de
naturaleza no covalente, se verá por qué estos complejos son necesariamente estéreo-específicos:
no se pueden formar más que si la molécula de enzima comporta un área
exactamente «complementaria» de la forma de la molécula de substrato. Se verá
también que, en el complejo, la molécula de substrato está necesariamente
colocada de manera muy rigurosa gracias a las múltiples interacciones que la
asocian al área receptora de la molécula de enzima.
Se verá, en fin, que según el número de
interacciones no covalentes que él pone en juego, la estabilidad de un complejo
no covalente podrá variar en una escala muy amplia. Es ésta una importante
propiedad de los complejos no covalentes: su estabilidad puede estar exactamente
adaptada a la función desempeñada. Los complejos enzima-substrato deben poder
hacerse y deshacerse muy rápidamente; es la condición de una alta actividad
catalítica. Estos complejos son, en efecto, fácil y muy rápidamente disociables.
Otros complejos, cuya función es permanente, adquieren una estabilidad del mismo
orden que la de una asociación covalente.
Sólo hemos discutido hasta el presente sobre la primera etapa de una reacción
enzimática: la formación del complejo estéreo-específico. La etapa catalítica en
sí, que sigue a la formación del complejo, no nos detendrá tanto, ya que no
plantea problemas tan profundamente significativos, desde un punto de vista
biológico, como la precedente. Se admite hoy en día que la catálisis enzimática
resulta de la acción inductora y polarizante de ciertos grupos químicos,
presentes en el «receptor específico» de la proteína. Especificidad aparte
(debida al posicionamiento muy preciso de la molécula de substrato respecto a
los grupos inductores), el efecto catalítico se explica por esquemas parecidos a
los que muestran la acción de los catalizadores no biológicos (como,
principalmente, los iones H+ y
OH−).
La formación del complejo estéreo-específico, como preludio al acto catalítico,
en sí puede pues ser considerada para el desempeño a la vez de dos funciones:
-
La elección exclusiva de un substrato, determinada por su
estructura estérica;
-
La presentación del substrato según una orientación precisa que
limita y especifica el efecto catalítico de los grupos inductores.
La noción de complejo estéreo-específico no covalente no se aplica sólo a las
enzimas ni incluso únicamente, como se verá, a las proteínas. Es de una
importancia central para la interpretación de todos los fenómenos de elección,
de discriminación electiva, que caracterizan a los seres vivos y les dan la
apariencia de escapar a la suerte que prevé el segundo principio. Es
interesante, a este respecto, considerar de nuevo el ejemplo de la fumarasa.
Si se realiza la aminación del ácido fumárico por los medios de la química
orgánica, se obtiene una mezcla de los dos isómeros ópticos del ácido aspártico.
La enzima, al contrario, cataliza exclusivamente la formación de ácido L-aspártico.
De este modo, aporta una información que corresponde con exactitud a una
elección binaria (ya que hay dos isómeros). Sé ve así, al nivel más elemental,
cómo la información estructural puede ser creada y distribuida en los seres
vivos. La enzima posee, desde luego, en la estructura de su receptor
estéreo-específico, la información correspondiente a esta elección. Pero la
energía necesaria para la amplificación de
esta información no viene de la enzima: para orientar la reacción exclusivamente
según uno de los dos caminos posibles, la enzima utiliza el potencial químico
constituido por la solución de ácido fumárico. Toda la actividad de síntesis de
las células, por compleja que sea, es, en último lugar, interpretable en los
mismos términos.
El demonio de Maxwell
Estos fenómenos, prodigiosos por su complejidad y su eficacia en la realización
de un programa fijado de antemano, imponen evidentemente la hipótesis de que son
guiados por el ejercicio de funciones de algún modo «cognitivas». Es una función
así la que Maxwell atribuyó a su demonio microscópico. Se recuerda que este
demonio, apostado en el orificio de comunicación entre dos recintos llenos de un
gas cualquiera, se suponía que maniobraba sin consumo de energía una trampa
ideal que le permitía prohibir el paso de ciertas moléculas de un recinto a
otro. El demonio podía pues «escoger» el no dejar pasar en un sentido más que a
las moléculas rápidas (de alta energía) y en el otro sólo las moléculas lentas
(de baja energía). El resultado era que, de los dos recintos inicialmente a la
misma temperatura, uno se calentaba mientras que el otro se enfriaba, todo ello
sin consumo aparente de energía. Para imaginamos lo que fue esta experiencia
habría que recordar el revuelo que causó en los medios científicos,
concretamente físicos: parecía en efecto que, por
el ejercicio de su función cognitiva,
el demonio tuviese el poder de violar el segundo principio. Y como esta función
cognitiva no parecía ni medible, ni incluso definible desde el punto de vista
físico, la «paradoja» de Maxwell parecía escapar a todo análisis en términos
operacionales.
La clave de la paradoja fue dada por Léon Brillouin, inspirándose en un trabajo
anterior de Szilard: demostró que el ejercicio de sus funciones cognitivas por
el demonio debía necesariamente consumir
una cierta cantidad de energía que, en el balance de la operación, compensaba
precisamente la disminución de entropía del sistema. En efecto, para que el
demonio cierre la trampa «con conocimiento de causa», es preciso que antes haya medido la
velocidad de cada partícula de gas. Luego, toda medida, es decir toda adquisición
de información,
supone una interacción, consumidora de energía.
Este célebre teorema es una de las fuentes de concepciones modernas relativas a
la equivalencia entre la información y la entropía negativa. Este teorema nos
interesa en nuestro caso por el hecho de que las enzimas ejercen precisamente, a
escala microscópica, una función creadora de orden. Pero esta creación de orden,
como hemos visto, no es gratuita; ella tiene lugar a expensas de un consumo de
potencial químico. Las enzimas, en definitiva, funcionan exactamente a la manera
del demonio de Maxwell corregido por Szilard y Brillouin, drenando el potencial
químico en las vías escogidas por el programa del que ellos son los ejecutantes.
Retengamos la noción esencial desarrollada en este capítulo: es gracias a su
capacidad de formar, con otras moléculas, complejos estéreo-específicos
y no covalentes,
por el que las proteínas ejercen sus funciones «demoníacas». Los capítulos
siguientes ilustrarán la importancia central de esta noción clave, que se
volverá a encontrar como interpretación última de las propiedades más
distintivas de los seres vivos.
Capítulo 4
Cibernética microscópica
Contenido:
Coherencia funcional de la maquinaria celular
Proteínas reguladoras y lógica de las regulaciones
Mecanismo de las interacciones alostéricas
Regulación de la síntesis de las enzimas
La noción de gratuidad
«Holismo» y reduccionismo
En virtud misma de su extrema especificidad, una enzima «clásica» (como las que
han sido tomadas como ejemplo en el capítulo precedente) constituye una unidad
funcional totalmente independiente. La función «cognitiva» de estos «demonios»
se reduce al reconocimiento de su substrato específico, excluyendo todo otro
cuerpo y todo acontecimiento que pueda producirse en la maquinaria química de la
célula.
Coherencia funcional de la maquinaria celular
La simple inspección de un esquema que resuma los conocimientos actuales sobre
el metabolismo celular bastaría para hacemos adivinar que si incluso, en cada
etapa, la enzima que tiene la carga cumple su tarea a la perfección, la suma
total de estas actividades no podría conducir más que a un caos si éstas no
estuviesen, de algún modo, sujetas las unas a las otras para formar un sistema
coherente. Así se tienen las pruebas más manifiestas de la eficacia extrema de
la maquinaria química de los seres vivos, de los más «simples» a los más
«complejos».
En los animales se conoce por descontado desde largo tiempo la existencia de
sistemas que aseguran la coordinación a gran escala de las performances del
organismo. Tales son las funciones del sistema nervioso y del sistema endocrino.
Estos sistemas aseguran la coordinación entre órganos o tejidos, es decir, en
definitiva, entre
células.
Que en el seno de cada célula una red cibernética casi tan compleja como ella
(si no más aún) asegura la coherencia funcional de la maquinaria química
intracelular, donde se centran los descubrimientos que datan, en su mayoría, de
los veinte si no de los cinco o diez últimos años.
Proteínas reguladoras y lógica de las regulaciones
Se está muy lejos aún de haber analizado en su integridad el sistema que
gobierna el metabolismo, el crecimiento y la división de las células más simples
que se conoce: las bacterias. Pero, gracias al análisis detallado de ciertas
partes de este sistema, se comprenden hoy bastante bien los principios de su
funcionamiento. Es de estos principios que discutiremos en el presente capitulo.
Veremos que las operaciones cibernéticas elementales están aseguradas por
proteínas especializadas, que juegan el papel de detectores e integradores de
información química.
Entre estas proteínas reguladoras, las mejor conocidas en la actualidad son
enzimas llamadas «alostéricas». Estas enzimas constituyen una clase particular,
en razón de las propiedades que las distinguen de las enzimas «clásicas». Como
estas últimas, las enzimas alostéricas reconocen y se asocian a un substrato
específico, y activan su conversión en productos. Pero además, estas enzimas
tienen la propiedad de reconocer electivamente uno o varios otros compuestos
cuya asociación (estéreo-específica) con la proteína tiene por efecto modificar,
es decir, según el caso, acrecentar
o inhibir su actividad con respecto al substrato.
La función reguladora, coordinadora, de las interacciones de este tipo (llamadas
interacciones alostéricas) está hoy en día probada por numerosos ejemplos. Se
pueden clasificar estas interacciones en un cierto número de «modos regulatorios»,
según las relaciones que existen entre la reacción considerada y el origen
metabólico de los «efectores alostéricos»
que la controlan. Los principales modos regulatorios son los siguientes (fig.
2).
Fig. 2. Diversos «modos regulatorios» asegurados por interacciones
alostéricas.
Las flechas en trazo grueso simbolizan reacciones produciendo cuerpos
intermediarios (A, B, etc.). La letra M representa el metabolito terminal,
resultado de la secuencia de reacciones. El trazo punteado indica el origen y el
punto de aplicación de un metabolito actuando como efector alostérico, inhibidor
o activador de una reacción (ver texto página anterior).
-
Inhibición retroactiva: la enzima que cataliza la primera reacción
de una secuencia que finaliza en un metabolito esencial (constituyente de las
proteínas o de los ácidos nucleicos, por ejemplo)[13] es
inhibida por el producto último de la secuencia. La concentración intracelular
de este metabolito gobierna pues la velocidad de su propia síntesis.
-
Activación retroactiva: la enzima es activada por un producto de
degradación del último metabolito. Este caso es frecuente en los metabolitos
cuyo potencial químico elevado constituye una moneda de cambio en el
metabolismo. Este modo de regulación contribuye pues a mantener a un nivel
prescrito el potencial químico disponible.
-
Activación en paralelo: la primera enzima de una secuencia
metabólica, que conduce a un metabolito esencial, es activada por un
metabolito sintetizado por una secuencia independiente y paralela. Este modo
de regulación contribuye a ajustar recíprocamente las concentraciones de
metabolitos pertenecientes a una misma familia y destinados a reunirse en una
de las clases de macromoléculas.
-
Activación por un precursor: la enzima es activada por un cuerpo
que es un precursor más o menos lejano de su substrato inmediato. Este modo de
regulación subordina en suma la «demanda» a la «oferta». Un caso particular,
extremadamente frecuente, de este modo regulatorio, es la activación de la
enzima por el substrato que desempeña a la vez su papel «clásico» y el de
efector alostérico con respecto a la enzima.
Es raro que una enzima alostérica esté sujeta sólo a uno de estos modos de
regulación. Por regla general estas enzimas están simultáneamente subordinadas a
varios efectores alostéricos, antagonistas o cooperativos. Una situación que se
encuentra frecuentemente es una regulación «ternaria» comprendiendo:
-
activación por el substrato (modo 4);
-
inhibición por el producto último de la secuencia (modo 1);
-
activación en paralelo por un metabolito de la misma familia que el último
producto (modo 3).
La enzima reconoce pues los tres efectores simultáneamente, «mide» sus
concentraciones relativas, y su actividad en todo instante representa la adición
de estas tres informaciones.
Con objeto de ilustrar el refinamiento de estos sistemas, se puede mencionar,
por ejemplo, los modos de regulación de las vías metabólicas «ramificadas» que
son numerosas (fig. 3). En ese caso, en general, no sólo las reacciones
iniciales, situadas en la bifurcación metabólica, son regularizadas por
inhibición retroactiva, sino que la reacción inicial de la rama común es
gobernada a la vez por los dos (o los varios) metabolitos finales.[14] El
peligro de bloqueo de la síntesis de uno de los metabolitos por un exceso del
otro es evitado, dependiendo de los casos, de dos formas diferentes:
-
sea asignando a esta reacción única dos enzimas alostéricas distintas,
cada una inhibida por uno de los metabolitos con exclusión del otro;
-
sea con una sola enzima que no es inhibida más que de manera «concertada»
por los dos metabolitos a la vez, pero no por uno solo de entre ellos.
Fig. 3. Regulación alostérica de las vías metabólicas ramificadas. Iguales
símbolos que en la figura 2 (ver texto).
Es preciso insistir en el hecho de que, substrato a parte, los efectores que
regulan la actividad de una enzima alostérica no participan en nada en la
reacción en sí. En general no forman con la enzima más que un complejo no
covalente, entera e instantáneamente reversible, de donde son liberados sin
ninguna modificación. El consumo de energía correspondiente a la interacción
reguladora es prácticamente nula: no representa más que una fracción ínfima del
potencial químico intracelular de los efectores. Al contrario, la reacción
catalítica gobernada por estas interacciones muy débiles puede implicar
transmisiones de energía relativamente considerables. Estos sistemas son, pues,
comparables a los que se emplean en los circuitos electrónicos de automoción,
donde la energía muy débil consumida por un relé puede desencadenar una
operación considerable, como, por ejemplo, el encendido de un cohete balístico.
* * * *
Igual que un relé electrónico puede estar subordinado simultáneamente a varios
potenciales eléctricos, igual, como se ha visto, una enzima alostérica lo está,
en general, a varios potenciales químicos. Pero la analogía va más lejos
todavía. Como se sabe, interesa generalmente que la respuesta de un relé
electrónico sea no
lineal respecto
a las variaciones de potencial que lo gobiernan. Se obtienen así efectos de
umbral que aseguran una regulación más precisa. Ello sucede igual en la mayoría
de las enzimas alostéricas. La gráfica, representando la variación de actividad
de una enzima de estas características en función de la concentración de un
efector (comprendido el substrato), es casi siempre «sigmoidal». En otros
términos, el efecto del ligando[15] crece más
deprisa que su concentración.
Esta propiedad es tanto más notable al ser característica de las enzimas
alostéricas. En las enzimas ordinarias, o «clásicas», el efecto crece siempre más
lentamente que
la concentración.
No sé cuál podría ser el peso mínimo de un relé electrónico presentando las
mismas propiedades lógicas que una enzima alostérica media (medida y suma de
tres o cuatro potenciales, solicitando una respuesta con efecto de umbral).
Supongamos que sea 10−2 gramos.
El peso de una molécula de una enzima alostérica capaz de las mismas performances es
del orden de 10−17gramos.
O sea, mil millones de veces menor que el relé electrónico. Este número
astronómico da una ligera idea de la «potencia cibernética» (es decir,
teleonómica) de la que puede disponer una célula provista de algunas centenas o
millares de especies de estos seres microscópicos, mucho más inteligentes aún
que el demonio de Maxwell-Szilard-Brillouin.
Mecanismo de las interacciones alostéricas
La cuestión es saber cómo estas performances complejas
son llevadas a cabo por este relé molecular que constituye una proteína
alostérica. Se admite hoy, sobre la base de un conjunto de hechos
experimentales, que las interacciones alostéricas son debidas a transiciones
discretas de estructura molecular de la misma proteína. Veremos en el próximo
capítulo que la estructura complicada y compacta de una proteína globular está
estabilizada por muy numerosas uniones no
covalentes que,
reunidas, cooperan al mantenimiento de la estructura Se concibe entonces que a
ciertas proteínas, dos (o varios) estados estructurales sean accesibles (igual
que ciertos cuerpos pueden existir en diferentes estados alotrópicos). Se
simbolizan a menudo los dos estados en cuestión, y la «transmisión alostérica»
que hace pasar la molécula reversiblemente del uno al otro de la manera
siguiente:

Sentado esto, se admite (y se demuestra directamente en los casos favorables)
que en razón de las estructuras estéticas diferentes
de los dos estados, las propiedades de reconocimiento estéreo-específicas de la
proteína son modificadas por la transición. Por ejemplo, en el estado «R», la
proteína podrá asociarse a un ligando α pero no a otro ligando β que será
reconocido (a exclusión de α) en el estado «T». La presencia de uno de los
ligandos tendrá pues por efecto estabilizar uno de los estados a expensas del
otro, y se ve que α y β serán antagonistas uno del otro, ya que sus asociaciones
respectivas con la proteína son mutuamente exclusivas. Supongamos ahora un
tercer ligando γ (que podría ser el substrato) asociándose exclusivamente con la
forma R, en una región de la molécula distinta a donde se fija α. Se ve que α y
γ cooperarán a la estabilización de la proteína en el estado activo (el que
reconoce el substrato). El ligando α y el substrato γ actuarán pues como
activadores, el ligando β, como inhibidor. La actividad de una población de
moléculas será proporcional a la fracción de ellas que estarán en el estado R,
fracción que depende evidentemente de la concentración relativa de los tres
ligandos, así como del valor del equilibrio intrínseco entre R y T. Es así que
la reacción catalítica se encontrará subordinada a los valores de estos tres
potenciales químicos.
Insistamos ahora sobre la noción más importante que implica este esquema: a
saber, que las interacciones cooperativas o antagonistas de los tres ligandos
son totalmente
indirectas. No hay, de hecho, interacciones entre los ligandos, sino
exclusivamente entre la proteína y cada uno de ellos separadamente.
Volveremos más adelante sobre esta noción fundamental, fuera de la cual parece
imposible comprender el origen y el desarrollo de los sistemas cibernéticos en
los seres vivos.[16]
A partir de este esquema de interacciones indirectas, es posible darse cuenta
igualmente del sutil perfeccionamiento que representa la respuesta «no lineal»
de la proteína a las variaciones de concentración de sus efectores. Todas las
proteínas alostéricas conocidas son en efecto «oligómeros», compuestos por la
asociación no covalente de subunidades (protómeros) químicamente idénticas, en
pequeño número (a menudo 2 o 4; más raramente 6, 8 o 12). Cada protómero lleva
un receptor para cada uno de los ligandos que la proteína reconoce. Por el hecho
de su asociación con uno o varios protómeros, la estructura estérica de cada uno
es parcialmente «constreñida» por sus vecinos. Pero la teoría, confirmada por la
experiencia de los cristalógrafos, demuestra que las proteínas oligoméricas
tienden a adoptar estructuras tales que todos los protómeros son geométricamente
equivalentes; los apremios que sufren son pues distribuidos simétricamente entre
los protómeros.
Tomemos ahora el caso más simple, el de un dímero: examinemos lo que lleva
consigo su disociación en dos monómeros; se ve que la ruptura de la asociación
va a permitir a los dos monómeros adoptar un estado «relajado», estructuralmente
diferente del «constreñido» que poseían en el estado asociado.
Diremos que el cambio de estado de los dos protómeros está «concertado». Es esta
concertación la que prueba la no linealidad de la respuesta; en efecto, la
estabilización por una molécula de ligando del estado disociado R en uno de los
monómeros prohíbe el retorno del otro al estado asociado, y sucede igual en
sentido inverso. El equilibrio entre los dos estados será una función cuadrática
de la concentración de los ligandos. Esto es, una función de potencia cuatro
para un tetrámero, y así consecutivamente[17].
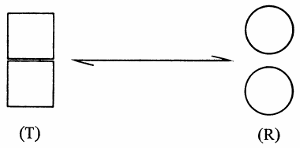
He tratado voluntaria y únicamente el modelo más posible, efectivamente
realizado por ciertos sistemas que podemos considerar como «primitivos». En los
sistemas reales, la disociación no es más que raramente completa: los protómeros
permanecen asociados en los dos estados, aunque de forma más laxa en uno de
ellos.
Numerosas variaciones son además posibles sobre este tema de base, pero lo
esencial era mostrar que mecanismos moleculares extremadamente simples en sí
mismos, permiten rendir cuenta de las propiedades «integrativas» de las
proteínas.
* * * *
Las enzimas alostéricas citadas hasta el presente constituyen a la vez una
unidad de función química y un elemento mediador de interacciones reguladoras.
Sus propiedades permiten comprender cómo el estado homeostático del metabolismo
celular es
conservado al máximo de eficacia y de coherencia.
Regulación de la síntesis de las enzimas
Se entiende por «metabolismo» esencialmente las transformaciones de las pequeñas
moléculas y la movilización del potencial químico. La química celular comprende
otro nivel de síntesis: el de las macromoléculas, ácidos nucleicos y proteínas
(comprendiendo principalmente las mismas enzimas). Se sabe desde hace tiempo que
a este nivel funcionan igualmente sistemas reguladores. El estudio es mucho más
difícil que el de las enzimas alostéricas, y de hecho uno solo de ellos ha
podido, hasta el presente, ser casi totalmente analizado. Lo tomaremos como
ejemplo.
Este sistema (llamado «sistema lactosa») gobierna la síntesis de tres proteínas
en la bacteria Escherichia
coli.
Una de estas proteínas (la galactósido-permeasa) permite a los galactósidos[18] penetrar
y acumularse en el interior de las células, cuya membrana, en ausencia de esta
proteína, es impermeable a estos azúcares. Una segunda proteína hidroliza los
β-galactósidos (ver capítulo 3). La función de la tercera proteína es sin duda
menor; no está del todo interpretada. Las dos primeras, por el contrario, son
simultáneamente indispensables para la utilización metabólica de la lactosa (y
otros galactósidos) por las bacterias.
Cuando éstas crecen en un medio desprovisto de galactósidos, las tres proteínas
son sintetizadas a un ritmo apenas mesurable, correspondiendo a una molécula
cinco generaciones por término medio. Casi inmediatamente (en dos minutos
aproximadamente) después de la adición de un galactósido (llamado en esta
circunstancia «inductor») al medio, el ritmo de síntesis de las tres proteínas
aumenta en un factor 1000, y se mantiene a este valor mientras que el inductor
está presente. Si el inductor se retira, el ritmo de síntesis vuelve a su valor
inicial en el espacio de dos a tres minutos.
Las conclusiones del análisis de este fenómeno, maravillosa y casi
milagrosamente teleonómico,[19] están
resumidas por el esquema de la figura 4.
Fig. 4. Regulación de la síntesis de las enzimas del «sistema lactosa». R:
Proteína-represor, en el estado asociado al galactósido inductor, representado
por un hexágono. T: Proteína-represor en el estado asociado al segmento operador
(o) del ADN. I: «Gen regulador» gobernando la síntesis del represor. p: Segmento
«promotor», punto de partida de la síntesis del RNA mensajero (mRNA). G1, G2,
G3: Gen de «estructura» que gobierna la síntesis de las tres proteínas del
sistema. P1, P2, P3. (Ver texto a continuación)
Se excluye aquí la discusión de la parte derecha del esquema que representa las
operaciones de síntesis del RNA «mensajero» y su «traducción» en secuencias
polipeptídicas. Retengamos solamente que el mensajero, pese a tener una vida muy
corta (del orden de algunos minutos), determina por su ritmo de síntesis el
ritmo de síntesis de las tres proteínas. Nos interesamos esencialmente por los
componentes del sistema regulador. Éstos comprenden:
—el gen «regulador» (i)
—la proteína-represor (R)
—el segmento «operador» (o) del ADN
—el segmento «promotor» (p) del ADN
—una molécula de galactósido inductor (βG)
El funcionamiento es el siguiente:
-
el gen regulador dirige la síntesis, a ritmo constante y muy débil, de la
proteína-represor;
-
el represor reconoce específicamente el segmento operador al cual se
asocia en un complejo muy estable (correspondiente a un ∆ F de cerca 15 Kcal);
-
en este estado la síntesis del mensajero (implicando la intervención de la
enzima RNA-polimerasa) está bloqueada, verosímilmente por simple impedimento
estérico, ya que el inicio de esta síntesis tiene lugar obligatoriamente al
nivel del promotor;
-
el represor reconoce igualmente los β-galactósidos, pero no se asocia
firmemente más que en estado libre; en presencia de β-galactósidos, por
consecuencia, el complejo operador-represor se disocia, ppermitiendo la
síntesis del mensajero, o sea de las proteínas.[20]
Es preciso subrayar que las dos interacciones del represor son reversibles y no
covalentes y que el inductor, particularmente, no es modificado por su
asociación con el represor. Así, la lógica de este sistema es de una extrema
simplicidad: el represor inactiva la transcripción; él es a su vez inactivado
por el inductor. De esta doble negación resulta un efecto positivo, una
«afirmación». Se puede observar que la lógica de esta negación de la negación no
es dialéctica: no desemboca en una proposición nueva, sino en la simple
reiteración de la proposición original, escrita en la estructura del ADN, en
conformidad con el código genético. La lógica de los sistemas biológicos de
regulación no obedece a la de Hegel, sino al álgebra de Boole, como la de las
calculadoras.
Se conoce hoy (en las bacterias) un gran número de sistemas análogos a éste.
Ninguno de ellos ha sido, hasta el presente, enteramente «desmontado». Parece
muy probable, sin embargo, que la lógica de algunos de estos sistemas sea más
complicada que la del sistema «lactosa», no comportando en especial y de forma
exclusiva interacciones negativas. Pero las nociones más generales y más
significantes que se pueden extraer del «sistema lactosa» son válidas también
para estos otros sistemas. Estas nociones son las siguientes:
-
El represor, desprovisto por él mismo de toda actividad, es un puro
mediador (transductor) de señales químicas.
-
El efecto del galactósido sobre la síntesis de la enzima es totalmente
indirecto, debido exclusivamente a las propiedades de reconocimiento del
represor y al hecho que dos estados, exclusivos uno del otro, le son
accesibles. Se trata pues claramente de una interacción alostérica en el
sentido del esquema general discutido más arriba.
-
No hay ninguna relación químicamente necesaria entre el hecho de
que la β-galactosidasa hidrolice los β-galactósidos, y el hecho de que su
biosíntesis sea inducida por los mismos cuerpos. Fisiológicamente útil,
«racional», esta relación es químicamente arbitraria. Se puede llamar
«gratuita».
La noción de gratuidad
Esta noción fundamental de gratuidad, es decir de independencia química entre la
misma función y la naturaleza de las señales químicas a las que está
subordinada, se aplica a las enzimas alostéricas. En este caso, una única
molécula de proteína desempeña a la vez la función catalítica específica y la
función reguladora. Pero, como se ha visto, las interacciones alostéricas son
indirectas, debidas exclusivamente a las propiedades diferenciales de
reconocimiento estéreo-específico de la proteína en los dos (o varios) estados
que le son accesibles. Entre el substrato de una enzima alostérica y los
ligandos que activan o inhiben su actividad, no existe ninguna relación químicamente
necesaria de
estructura o de reactividad. La especificidad de las interacciones es, en
definitiva, independiente de la estructura de los ligandos: es debida
enteramente a la de la proteína en los diversos estados que le son accesibles,
estructura a su vez, libremente, arbitrariamente dictada por
la de un gen.
Así resulta, y éste es el punto fundamental, que por lo que atañe a la
regulación por el intermediario de una proteína alostérica todo
es posible.
Una proteína alostérica debe ser considerada como un producto especializado de
«ingeniería» molecular, permitiendo a una interacción, positiva o negativa,
establecerse entre cuerpos desprovistos de afinidad química y así subordinar una
reacción cualquiera a la intervención de compuestos químicamente extraños e
indiferentes a esta reacción. El principio operatorio de las interacciones
alostéricas autoriza pues una entera libertad en la «elección» de los
subordinados que, escapando a todo apremio químico, podrán obedecer
exclusivamente a los apremios fisiológicos en virtud de los que serán
seleccionados según el aumento de coherencia y de eficacia que confieren a la
célula o al organismo. Es en definitiva la gratuidad misma
de estos sistemas que, abriendo a la evolución molecular un campo prácticamente
infinito de exploración y de experiencias, le ha permitido construir la inmensa
red de interconexiones cibernéticas que hacen de un organismo una unidad
funcional autónoma, de la que las performances parecen
transcender las leyes de la química, sino incluso escapar de ellas.
De hecho, como se ha visto, cuando estas performances son
analizadas a escala microscópica, molecular, aparecen enteramente interpretables
en términos de interacciones químicas específicas, electivamente aseguradas,
libremente escogidas y organizadas por proteínas reguladoras; y es en la
estructura de estas moléculas donde es preciso ver la fuente última de la
autonomía, o más exactamente de la autodeterminación que caracteriza a los seres
vivos en sus performances.
Los sistemas que hemos estudiado hasta aquí son de los que coordinan la
actividad de la célula y hacen de ella una unidad funcional. En los organismos
pluricelulares, sistemas especializados aseguran la coordinación entre células,
tejidos u órganos: se trata no sólo del sistema nervioso y del sistema
endocrino, sino también de las interacciones directas entre células. No abordaré
aquí el análisis del funcionamiento de estos sistemas que escapa aún, casi
enteramente, a la descripción microscópica. Admitiremos, sin embargo, la
hipótesis que, en estos sistemas, las interacciones moleculares que aseguran la
transmisión y la interpretación de las señales químicas son debidas a proteínas
dotadas de propiedades de reconocimiento estéreo-específicas diferenciales, a
las que se aplica el principio esencial de gratuidad química, tal como se
desprende del estudio de las interacciones alostéricas propiamente dichas.
«Holismo» y reduccionismo
Cabría quizá, para concluir este capítulo, volver de nuevo sobre la antigua
disputa entre «reduccionistas» y «organicistas». Se sabe que ciertas escuelas
del pensamiento (todas más o menos conscientemente o confusamente influenciadas
por Hegel) niegan el valor del acceso analítico cuando
se trata de sistemas tan complejos como los seres vivos. Según estas escuelas
(«organicistas» u «holistas») que, como el fénix, renacen en cada generación,[21] la
actitud analítica, calificada de «reduccionista», sería completamente estéril,
como pretendiendo recoger pura y simplemente las propiedades de una organización
muy compleja en la «suma» de sus partes. Es una querella estúpida, que atestigua
solamente, en los «holistas», un profundo desconocimiento del método científico
y del papel esencial que en él juega el análisis. ¿Podemos acaso concebir que un
ingeniero marciano, queriendo interpretar el funcionamiento de una calculadora
terrestre, pueda llegar a algún resultado si rehúsa, por principio, disecar los
componentes electrónicos de base que efectúan las operaciones de álgebra
proposicional? Si hay un dominio de la biología molecular que ilustre más que
otros la esterilidad de las tesis organicistas por oposición al poderío del
método científico, es desde luego el estudio de esta cibernética microscópica
brevemente entrevisto en el curso de este capítulo.
El análisis de las interacciones alostéricas muestra, para empezar, que las performances teleonómicas
no son el atributo exclusivo de sistemas complejos, de componentes múltiples, ya
que una molécula
de proteína se muestra ya capaz, no sólo de activar electivamente una reacción,
sino de regular su actividad en función de varias informaciones
químicas.
Vemos, en segundo lugar, gracias a la noción de gratuidad, cómo y por qué estas
interacciones reguladoras moleculares, escapando a los apremios químicos, han
podido ser selectivamente escogidas en razón exclusivamente de su participación
en la coherencia del sistema.
El estudio de estos sistemas microscópicos nos revela, en fin, que la
complejidad, la riqueza y la potencia de la red cibernética, en los seres vivos,
sobrepasan con mucho lo que el estudio de las solas performances globales
de los organismos podría jamás dejar entrever. E incluso cuando esos análisis
están lejos aún de suministrar una descripción completa del sistema cibernético
de la célula más simple, revelan que todas las actividades, sin excepción, que
concurren al crecimiento y a la multiplicación de esta célula están,
directamente o no, subordinadas unas a otras.
Es sobre estas bases, y no sobre la de una vaga «teoría general de los
sistemas»,[22] que
nos es posible comprender en qué sentido, muy real, el organismo trasciende, en
efecto, aunque observándolas, las leyes físicas para no ser más que persecución
y cumplimiento de su propio proyecto.
Capítulo 5
Ontogénesis molecular
Contenido:
La asociación espontánea de las subunidades en las proteínas
oligoméricas
Estructuración espontánea de partículas complejas
Morfogénesis microscópica y morfogénesis macroscópica
Formación de las estructuras globulares
La falsa paradoja del «enriquecimiento» epigenético
La ultima ratio de las estructuras teleonómicas
La interpretación del mensaje
Los seres vivos, tanto por sus estructuras macroscópicas como por sus funciones,
son, lo hemos visto, estrechamente comparables a las máquinas. Difieren
radicalmente, por el contrario, por su modo de construcción. Una máquina, un
artefacto cualquiera, debe su estructura macroscópica a la acción de fuerzas
exteriores, de útiles que actúan sobre una materia para imponerle su forma. Es
el cincel del escultor el que arranca del mármol las formas de Afrodita; pero la
diosa, no su imagen, nació de la espuma de las olas (fecundadas por el órgano
sangrante de Urano) de donde su cuerpo creció de sí mismo, por sí mismo.
Querría, en este capítulo, mostrar que este proceso de morfogénesis espontánea y
autónoma reposa, en último lugar, en las propiedades de reconocimiento
estéreo-específico de las proteínas, que es pues de orden microscópico, antes de
manifestarse en estructuras macroscópicas. Buscaremos, en conclusión, en las
estructuras primarias de las proteínas el «secreto» de estas propiedades
cognitivas que convierten a los demonios de Maxwell en animadores y
constructores de los sistemas vivientes.
Es preciso subrayar, en primer lugar, que los problemas que ahora abordamos, los
de la mecánica del desarrollo, plantean aún a la biología profundos enigmas.
Porque si la embriología ha aportado admirables descripciones del desarrollo, se
está aún lejos de saber analizar la ontogénesis de las estructuras macroscópicas
en términos de interacciones microscópicas. Por el contrario, la construcción de
ciertos edificios moleculares está hoy bastante bien comprendida y me gustaría
mostrar que se trata de un verdadero proceso «de ontogénesis molecular» donde se
revela la esencia física del fenómeno.
He tenido ya ocasión de recordar que las proteínas globulares se presentan a
menudo bajo forma de agregados conteniendo un número finito de subunidades
químicamente idénticas. El número de las subunidades constituyentes, siendo
generalmente pequeño, permite llamar a estas proteínas «oligómeros». En éstos
oligómeros, las subunidades (protómeros) están asociadas exclusivamente por
uniones no covalentes. Además, como ya se ha visto, el orden de los protómeros
en el seno de una molécula oligomérica es tal que cada uno de ellos es
equivalente, geométricamente, a cada uno de los otros. Esto entraña
necesariamente que cada protómero pueda ser convertido en cualquiera de los
otros por una operación de simetría, de hecho por una rotación. Se demuestra
fácilmente que los oligómeros así constituidos poseen los elementos de simetría
de uno de los grupos puntuales de rotación.
Estas moléculas constituyen, pues, verdaderos cristales microscópicos, pero
pertenecientes a una clase particular que yo llamaría la de los «cristales
cerrados» porque, contrariamente a los cristales propiamente dichos (construidos
según uno de los grupos de espacio), no pueden crecer sin adquirir nuevos
elementos de simetría, todo y perdiendo (en general) algunos de los que poseían.
La asociación espontánea de las subunidades en las proteínas
oligoméricas
Hemos visto ya, en fin, que ciertas propiedades funcionales de estas proteínas
están asociadas a su estado oligomérico y a su estructura simétrica. La
construcción de estos edificios microscópicos plantea pues un problema
biológicamente significante, al mismo tiempo que físicamente interesante.
Como los protómeros, en una molécula oligomérica, no están asociados más que por
uniones no covalentes, es a menudo posible, por tratamientos muy moderados (no
implicando, por ejemplo, la presencia de agentes químicos agresivos o el alcance
de temperaturas elevadas), disociarlos en unidades monoméricas. En este estado,
la proteína ha perdido en general todas sus propiedades funcionales, catalíticas
o reguladoras. Luego, y éste es el punto importante, cuando las condiciones
iniciales «normales» son restituidas (por eliminación del agente disociante), se
constata en general que los agregados oligoméricos se reforman espontáneamente,
con restauración completa del estado «nativo»: mismo número de protómeros, misma
simetría, acompañada de la reaparición integral de las propiedades funcionales.
Además, la re asociación de subunidades perteneciendo a una misma especie
proteínica no se produce sólo en una solución conteniendo esta única proteína.
Ella tiene lugar, igualmente, en mezclas complejas conteniendo centenas, si no
millares, de otras proteínas. Prueba que hay, una vez más, un proceso de
reconocimiento de una extrema especificidad, evidentemente debido a la formación
de complejos estéticos no covalentes asociando los protómeros entre sí. Proceso
que es legítimo considerar como epigenético,[23] ya
que a partir de una solución de moléculas monoméricas, desprovistas de toda
simetría, han aparecido moléculas mayores y de un grado de orden superior que,
del mismo golpe, han adquirido propiedades funcionales, antes totalmente
ausentes.
Lo esencial, para lo que aquí nos interesa, es el carácter
espontáneo de
este proceso de epigénesis molecular. Espontáneo en dos sentidos.
-
El potencial químico necesario para la formación de los oligómeros no ha
de ser inyectado en el sistema: se debe considerar que está presente en la
solución de monómeros.
-
Termodinámicamente espontáneo, el proceso lo es de la misma manera
cinéticamente: ningún catalizador es requerido para activarlo. Esto gracias,
evidentemente, al hecho de que las uniones empleadas son no covalentes. Hemos
ya subrayado la extrema importancia del hecho de que la formación, como la
ruptura, de tales uniones no pone en juego más que energías de activación casi
nulas.
Estructuración espontánea de partículas complejas
Un fenómeno así es estrechamente comparable a la formación de cristales
moleculares a partir de una solución de las moléculas constituyentes. Aquí
también hay constitución espontánea de orden, por asociación entre ellas, de
moléculas pertenecientes a una misma especie química. La analogía es tanto más
evidente ya que, en los dos casos, se ven formarse estructuras ordenadas según
reglas geométricas simples y repetitivas. Pero se ha podido mostrar
recientemente que ciertos orgánulos celulares de estructura mucho más compleja
son igualmente los productos de un ensamblaje espontáneo. Es el caso de las
partículas llamadas ribosomas, que son componentes esenciales del mecanismo de
traducción del código genético, es decir de la síntesis de las proteínas. Estas
partículas, cuyo peso molecular alcanza 10−6,
están constituidas por el ensamblaje de unas treinta proteínas distintas así
como de tres tipos diferentes de ácidos nucleicos. Aunque la disposición exacta
de estos diferentes constituyentes en el seno de un ribosoma no sea conocida, es
cierto que la organización es extremadamente precisa y que la actividad
funcional de la partícula depende de ella. Por lo tanto, a partir de los
constituyentes disociados de los ribosomas, se asiste a la reconstitución
espontánea, in
vitro,
de partículas de igual composición, de igual peso molecular, poseyendo la misma
actividad funcional que el material «nativo» inicial.[24]
Sin duda, sin embargo, el ejemplo más espectacular que se conoce hoy en día de
construcción espontánea de un edificio molecular complejo, es el de ciertos
bacteriófagos.[25] La
estructura complicada y muy precisa del bacteriófago T4 corresponde a la función
de esta partícula, que no es solamente proteger al genoma (es decir el ADN) del
virus, sino de adherirse a la pared de la célula huésped para inyectar, a la
manera de una jeringa, su contenido de ADN. Las diferentes piezas de esta
maquinaria microscópica de precisión pueden ser obtenidas separadamente a partir
de diferentes mutantes del virus. Mezcladas in
vitro,
se ensamblan espontáneamente para
reconstituir partículas idénticas a las normales, y plenamente capaces de
ejercer su función de jeringa de ADN.[26]
Todas estas observaciones son relativamente recientes, y se pueden esperar
progresos importantes en este dominio de la investigación, desembocando en la
reconstitución in
vitro de
orgánulos celulares cada vez más complejos, tales como mitocondrias o membranas,
por ejemplo. Los casos a los que hemos pasado aquí revista bastan sin embargo
para ilustrar el proceso por el que estructuras complejas, a las que están
ligadas propiedades funcionales, son construidas por el ensamblaje
estéreo-específico, espontáneo,
de sus constituyentes proteínicos. Hay «aparición» de orden, diferenciación
estructural, adquisición de funciones a partir de una mezcla desordenada de
moléculas individualmente desprovistas de toda actividad, de toda propiedad
funcional intrínseca que no sea reconocer los compañeros con los que van a
constituir la estructura. Y si no se puede, para los ribosomas o los
bacteriófagos, hablar de cristalización, ya que estas partículas son de un grado
de complejidad, es decir de orden, muy superior al que caracteriza un cristal,
no deja de ser cierto que, en el fondo, las interacciones químicas puestas en
juego son de la misma naturaleza que las que construyen un cristal molecular.
Como en un cristal, es la estructura misma de las moléculas ensambladas lo que
constituye la fuente de «información» para la construcción del conjunto. La
esencia de estos procesos epigenéticos consiste pues en que la organización de
conjunto de un edificio multimolecular complejo está contenido en potencia en la
estructura de sus constituyentes, pero no se revela, ni deviene actual más
que por su ensamblaje.
Este análisis se ve reducido a una disputa verbal, desprovista de todo interés:
la antigua querella de los preformacionistas y de los epigenistas. La estructura
acabada no estaba en ninguna parte, como tal, preformada. Pero el plan de la
estructura estaba presente en sus mismos constituyentes. Ella puede pues
realizarse de forma autónoma y espontánea, sin intervención exterior, sin
inyección de nueva información. La información estaba presente, pero sin
expresar, en los constituyentes. La construcción epigenética de una estructura
no es una creación,
es una revelación.
Morfogénesis microscópica y morfogénesis macroscópica
Que esta concepción, directamente fundamentada sobre el estudio de la formación
de edificios microscópicos, pueda y deba igualmente rendir cuenta de la
epigénesis de las estructuras macroscópicas (tejidos, órganos, miembros, etc.),
es algo que los modernos biólogos no dudan, aun reconociendo que se trata de una
extrapolación a la que faltan todavía las verificaciones directas. Estos
problemas se plantean en efecto a otra escala, no sólo en dimensiones, sino en
complejidad. Las interacciones constructivas más importantes en esta escala
tienen lugar no entre componentes moleculares, sino entre células. Se ha podido
ver que células aisladas de un mismo tejido son efectivamente capaces de
reconocerse entre ellas, diferencialmente, y de juntarse. Sin embargo, se ignora
todavía cuáles son los componentes o las estructuras que identifican las células
unas respecto a otras. Todo lleva a creer que se trata de estructuras
características de las membranas celulares. Pero no se sabe si estos elementos
de reconocimiento son estructuras moleculares tomadas individualmente o redes
multimoleculares de superficie.[27] Sea
lo que sea, e incluso si se trata de redes que no están constituidas
exclusivamente por proteínas, la estructura de tales redes tendrá
necesariamente, en último caso, que ser determinada por las propiedades de
reconocimiento de sus constituyentes proteínicos, así como por las de las
enzimas responsables de la biosíntesis de los otros componentes de la red
(polisacáridos o lípidos, por ejemplo).
Por tanto, es posible que las propiedades «cognitivas» de las células no sean la
manifestación directa de las facultades discriminativas de algunas proteínas,
pero no expresan estas facultades más que por vías muy apartadas. Lo que no
impide que la construcción de un tejido o la diferenciación de un órgano,
fenómeno macroscópico, deban ser considerados como la resultante integrada de
interacciones microscópicas múltiples debidas a proteínas, y reposando sobre sus
propiedades de reconocimiento estéreo-específico, por formación espontánea de
complejos no covalentes.
Mas es preciso reconocer que esta «reducción a lo microscópico» de los fenómenos
de la morfogénesis no constituye, por el momento, una verdadera teoría de estos
fenómenos. Se trata más bien de una posición de principio que especifica
solamente los términos en los que una teoría así debería ser formulada para que
se pueda considerar como aportando más que una simple descripción
fenomenológica. Este principio define el fin a alcanzar, pero no ilumina más que
débilmente la vía a seguir para conseguirlo. Piénsese en el formidable problema
que representa la interpretación, a escala molecular, del desarrollo de un
aparato tan complejo como el sistema nervioso central, en el que miles de
millones de interconexiones específicas entre células deben ser realizadas,
algunas a distancias relativamente considerables.
Este problema de las influencias, de las orientaciones a distancia, es sin duda
el más difícil y el más importante de la embriología. Los embriólogos, para
explicar con facilidad los fenómenos de regeneración, han introducido la noción
de «campo morfogenético» o de «gradiente». Noción que parece al principio
sobrepasar de lejos la de interacción estéreo-específica a escala de algunos
Angstrom. Resulta, sin embargo, que esta última es la única en presentar un
sentido físico preciso, y que no es del todo inconcebible que tales
interacciones, multiplicadas y repetidas progresivamente, puedan crear o definir
una organización a escala milimétrica, o centimétrica por ejemplo. Es en este
sentido que se orienta la embriología moderna. Es bastante verosímil que la
noción de interacciones estéreo-específicas puramente estáticas resulte
insuficiente para la interpretación del «campo» o de los gradientes
morfogenéticos. Será preciso enriquecerla de hipótesis cinéticas, análogas
quizás a las que permiten la interpretación de las interacciones alostéricas.
Pero yo sigo convencido, por mi parte, que sólo las propiedades asociativas
estéreo-específicas de las proteínas podrán, en último lugar, dar la clave de
estos fenómenos.
Estructura primaria y estructura globular de las proteínas
Cuando se analizan las funciones catalíticas o reguladoras o epigenéticas de las
proteínas, se es llevado a reconocer que todas, y ante todo, reposan sobre las
propiedades asociativas estéreo-específicas de estas moléculas.
Según la concepción expuesta en este capítulo, como en los dos precedentes,
todas las performances y
todas las estructuras teleonómicas de los seres vivos son, al menos en
principio, analizables en estos términos. Si esta concepción es adecuada —y no
hay razón para dudar que lo sea— resta pues, para resolver la paradoja de la
teleonomía, explicar el modo de formación y los mecanismos de evolución de las
estructuras asociativas estéreo-específicas de las proteínas. No examinaré aquí
más que el modo de formación de estas estructuras, reservando la cuestión de su
evolución para los próximos capítulos. Espero mostrar que el análisis detallado
de estas estructuras moleculares que, en suma, guardan el «secreto» último de la
teleonomía, conduce a conclusiones profundamente significantes.
Para empezar, es necesario recordar que la estructura en el espacio de una
proteína globular (cf. Apéndice, 1. Estructura
de las proteínas)
está determinada por dos tipos de uniones químicas.
-
La estructura llamada «primaria» está constituida por una secuencia
topológicamente lineal de radicales de aminoácidos asociados por uniones
covalentes. Estas uniones definen pues una estructura fibrosa, extremadamente
flexible, y capaz de tomar, en teoría, una infinidad de conformaciones.
-
Pero la conformación llamada «nativa» de una proteína globular, está
además estabilizada por un gran número de interacciones no covalentes que
asocian entre ellas los radicales de aminoácidos repartidos a lo largo de la
secuencia covalente topológicamente lineal. De ello resulta que la fibra
polipeptídica está plegada sobre ella misma de manera muy compleja, en una
pelota pseudo globular, compacta. Son, en definitiva, estos pliegues complejos
los que determinan la estructura en el espacio de la molécula, comprendida la
forma precisa de las áreas de asociación estéreo-específica por la que la
molécula ejerce su función de reconocimiento. Es, pues, como se ve, la suma, o
mejor la cooperación de un gran número de interacciones no covalentes que
estabilizan la estructura funcional, lo que permite a la proteína formar
electivamente complejos estéreo-específicos (igualmente no covalentes) con
otras moléculas.
La cuestión que aquí nos interesa es la ontogénesis, el modo de formación de
esta conformación particular, única, a la que está ligada la función cognitiva
de una proteína. Se ha podido creer, durante mucho tiempo, que en razón de la
complejidad misma de estas estructuras y del hecho de que ellas están
estabilizadas por interacciones no covalentes, individualmente muy lábiles, un
gran número de conformaciones distintas serian accesibles a una misma fibra
polipeptídica. Pero todo un conjunto de observaciones debía mostrar que de hecho
una misma especie química (definida por la estructura primaría) no existe en el
estado nativo, en las condiciones fisiológicamente normales, más que en una sola
conformación (o todo lo más, en un pequeñísimo número de estados distintos, poco
distantes unos de otros, como es el caso de las proteínas alostéricas).
Conformación muy precisamente definida, como lo prueba el hecho de que los
cristales de proteínas dan excelentes imágenes de difracciones de rayos X, lo
que significa que la posición de la gran mayoría de los millares de átomos que
componen una molécula está fijada con un error de sólo algunas fracciones de
Angstrom. Señalemos, además, que esta uniformidad, como esta precisión de
estructura, son la condición misma de la especificidad de asociación, propiedad
biológicamente esencial de las proteínas globulares.
Formación de las estructuras globulares
El mecanismo de formación de estas estructuras está, en el presente, bastante
bien conocido en su principio. Se sabe en efecto:
-
Que el determinismo genético de las estructuras de proteínas especifica
exclusivamente la secuencia de los radicales aminoácidos que corresponden
a una proteína dada;
-
Que la fibra polipeptídica así sintetizada se pliega espontáneamente y de
forma autónoma para concluir en la conformación pseudo globular,
funcional.
Así, entre los millares de conformaciones plegadas, en principio accesibles a la
fibra polipeptídica, una sola es de hecho escogida y realizada. Se trata, como
se ve, de un verdadero proceso epigenético, al nivel más simple posible: el de
una macromolécula aislada. A la fibra desplegada, le son accesibles millares de
conformaciones. Ella está, por otra parte, desprovista de toda actividad
biológica, especialmente de toda capacidad de reconocimiento estéreo-específico.
En la forma plegada, por el contrario, un solo estado le es accesible, que
corresponde por consecuencia a un nivel de orden muy superior. Es en este
estado, exclusivamente, que es conferida la actividad funcional.
La explicación de este pequeño milagro de epigénesis molecular es relativamente
simple en su principio.
-
En el medio fisiológicamente normal, es decir, en fase acuosa, las formas
plegadas de la proteína son termodinámicamente más estables que las formas
desplegadas. La razón de esta diferencia de estabilidad es muy interesante; es
importante precisarla. Entre los radicales aminoácidos que constituyen la
secuencia, la mitad aproximadamente son «hidrófobos», es decir, se comportan
como el aceite en el agua, tienden a juntarse liberando las moléculas de agua
inmovilizadas a su contacto. Por este hecho, la proteína toma una estructura
compacta, inmovilizando, por contacto reciproco, los radicales que componen la
fibra, de donde, para las proteínas, una ventaja de orden (o de entropía
negativa) compensada por la expulsión de moléculas de agua, que, liberadas,
van a acrecentar el desorden, es decir, la entropía del sistema.
-
Entre las diferentes estructuras plegadas accesibles a una secuencia
polipeptídica dada, una sola, o un pequeñísimo número de ellas, permiten la
realización de la estructura más compacta posible. Esta estructura será pues
privilegiada a expensas de todas las demás. Digamos, simplificando un poco,
que será «escogida» la que correspondía a la expulsión de un máximo de
moléculas de agua. Evidentemente, es de la posición relativa, es decir de la
secuencia de los radicales aminoácidos en la fibra (comenzando por los
radicales hidrófobos) que dependerán las diferentes posibilidades de
realización de estructuras compactas. La conformación globular particular en
una proteína dada y de la que depende su actividad funcional será pues de
hecho impuesta por la secuencia de los radicales en la fibra. Sin
embargo, y éste es el punto importante, la cantidad de información que sería
necesaria para especificar enteramente la estructura tridimensional de una
proteína es mucho más grande que la información definida
por la misma secuencia. Por ejemplo, para un polipéptido de cien aminoácidos,
la información (H) necesaria para la definición de la secuencia,
correspondería a unos 2.000 bits (H = log2 20100),
mientras que para definir la estructura tridimensional sería preciso, a este
número, añadirle aún una gran cantidad de información, difícilmente calculable
además (digamos 1.000 a 2.000 bits, al menos).
-
La falsa paradoja del «enriquecimiento» epigenético
Se puede ver, pues, una contradicción en el hecho de decir que el genoma «define
enteramente» la función de una proteína, mientras que esta función está ligada a
una estructura tridimensional cuyo contenido informativo es más
rico que
la contribución directamente aportada a esta estructura por el determinismo
genético. Esta contradicción no ha dejado de ser realzada por ciertos críticos
de la teoría biológica moderna. Especialmente Elsässer, quien ve precisamente en
el desarrollo epigenético de las estructuras (macroscópicas) de los seres vivos
un fenómeno físicamente inexplicable, en razón del «enriquecimiento sin causa»
que él parece testimoniar.
Esta objeción desaparece cuando se examinan con detalle los mecanismos de la
epigénesis molecular, el enriquecimiento de información, correspondiente a la
formación de la estructura tridimensional, proviene de lo que la información
genética (representada por la secuencia) expresa de hecho en condiciones
iniciales bien definidas (en fase acuosa, entre ciertos límites, estrechos, de
temperaturas, composición iónica, etc.), tales que, entre todas las estructuras
posibles, una sola de ellas es de hecho realizable. Las condiciones iniciales,
en consecuencia, contribuyen a la información finalmente encerrada en la
estructura globular, sin por otra parte especificarla, sino solamente eliminando
las demás estructuras posibles, proponiendo así, o más bien imponiendo, una
interpretación unívoca de un mensaje a
priori parcialmente
equivoco.
* * * *
En el proceso de estructuración de una proteína globular, se puede ver, pues, a
la vez, la imagen microscópica y la fuente del desarrollo epigenético autónomo
del organismo. Desarrollo en el que se pueden reconocer varias etapas o niveles
sucesivos.
-
Pliegue de las secuencias polipeptídicas para dar las estructuras
globulares, provistas de las propiedades asociativas estéreo-específicas.
-
Interacciones asociativas entre proteínas (o entre proteínas y algunos
otros constituyentes) para formar los orgánulos celulares.
-
Interacciones entre células, para constituir tejidos y órganos.
-
En todas estas etapas, coordinación y diferenciación de las actividades
químicas, por interacciones de tipo alostérico.
En cada una de estas etapas aparecen estructuras de orden superior y funciones
nuevas que, resultando de las interacciones espontáneas entre productos de la
etapa precedente, revelan, como en un fuego de artificio de varios pisos, las
potencialidades latentes de los niveles anteriores. Todo el determinismo del
fenómeno encuentra su fuente, en definitiva, en la información genética
representada por la suma de las secuencias polipeptídicas interpretadas, o más
exactamente filtradas, por las condiciones iniciales.
La ultima ratio de las estructuras teleonómicas
La ultima
ratio de
todas las estructuras y performances teleonómicas
de los seres vivos está, pues, encerrada en las secuencias de radicales de las
fibras polipeptídicas, «embriones» de estos demonios biológicos de Maxwell que
son las proteínas globulares. En un sentido, muy real, es a este nivel de
organización química donde yace, si es que hay uno, el secreto de la vida. Y
sabiendo no sólo describir estas secuencias, sino enunciar la ley de ensamblaje
a la que ellas obedecen, se podría decir que el secreto ha sido penetrado, la ultima
ratio descubierta.
La primera secuencia completa de una proteína globular fue descrita en 1952 por
Sanger. Esto fue a la vez una revelación y una decepción. En esta secuencia en
la que se sabía definir la estructura, o sea las propiedades electivas de una
proteína funcional (la insulina), ninguna regularidad, ninguna singularidad,
ninguna restricción se revelaba. Aunque, sin embargo, se puede esperar que, a
medida que se acumulen documentos de este tipo, algunas leyes generales de
ensamblaje, así como ciertas correlaciones funcionales, se harán más claras. Se
conocen hoy en día centenares de secuencias, correspondientes a distintas
proteínas, extraídas de los organismos más diversos. De estas secuencias, y de
su comparación sistemática ayudada por los modernos medios de análisis y de
cálculo, se puede hoy deducir la ley general: la del azar. Para ser precisos:
estas estructuras están «al azar» en el sentido que, conociendo exactamente el
orden de 199 residuos en una proteína que comprende 200, es imposible formular
ninguna regla, teórica o empírica, que permita prever la naturaleza del único
residuo no identificado aún por el análisis.
Decir de la secuencia de los aminoácidos en un polipéptido que está «al azar» no
agrega nada, hay que insistir sobre ello, a una declaración de ignorancia; sino
que expresa una constatación de hecho: por ejemplo, la frecuencia media con la
cual el residuo está seguido de uno determinado en los polipéptidos es igual al producto de
las frecuencias medias de cada uno de los dos residuos en las proteínas en
general. Se puede ilustrar esto de otra forma. Supongamos un juego de cartas en
el que cada una lleva el nombre de un aminoácido. Sea un paquete de doscientas
cartas en el que la proporción media de
cada aminoácido sería respetada. Después de haber barajado las cartas se
obtendrían secuencias al azar, que nada permitiría distinguir de las secuencias
efectivamente observadas en los polipéptidos naturales.
Pero si, en este sentido, toda estructura primaria de proteína nos aparece como
el puro producto de una elección hecha al azar, en cada eslabón, entre los
residuos disponibles, por el contrario en otro sentido, también muy
significativo, es preciso reconocer que esta secuencia actual no
ha sido sintetizada en modo alguno al azar, ya que este mismo orden está
reproducido, prácticamente sin error, en todas las moléculas de la proteína
considerada. No siendo así, sería imposible, de hecho, establecer por el
análisis químico la secuencia de una población de moléculas.
Así, pues, es preciso admitir, que la secuencia «al azar» de cada proteína está
de hecho reproducida, millares o millones de veces, en cada organismo, en cada
célula, en cada generación, por un mecanismo de alta fidelidad que asegura la
invariancia de las estructuras.
La interpretación del mensaje
Se conocen hoy no sólo el principio, sino la mayoría de los componentes de este
mecanismo. Se volverá sobre ello en un próximo capítulo. No es necesario conocer
los detalles de este mecanismo para comprender la significación profunda del
misterioso mensaje que constituye la secuencia de los radicales en una fibra
polipeptídica. Mensaje que, según todos los criterios posibles, parece haber
sido escrito al azar. Mensaje, sin embargo, cargado de un sentido que se revela
en las interacciones discriminativas, funcionales, directamente teleonómicas, de
la estructura globular, traducción a tres dimensiones de la secuencia lineal.
Una proteína globular es ya, a escala molecular, una verdadera máquina por sus
propiedades funcionales, pero no, lo vemos ahora, por su estructura fundamental,
donde nada se discierne más que el juego de combinaciones ciegas. Azar captado,
conservado, reproducido por la maquinaria de la invariancia y así convertido en
orden, regla, necesidad. De un juego totalmente ciego,
todo, por definición, puede salir, incluida la misma visión. En la ontogénesis
de una proteína funcional, el origen y la filiación de la biosfera entera se
reflejan, y la fuente última del proyecto que los seres vivos representan,
persiguen y cumplen, se revela en este mensaje, en este texto preciso, fiel,
pero esencialmente indescifrable que constituye la estructura primaría.
Indescifrable, ya que antes de expresar la función fisiológicamente necesaria
que él cumple espontáneamente, no revela en su estructura más que el azar de su
origen. Pero tal es, justamente, el sentido más profundo de este mensaje que nos
llega del fondo de las edades.
Capítulo 6
Invariancia y perturbaciones
Contenido:
Platón y Heráclito
Las invariantes anatómicas
Las invariantes químicas
El ADN como invariante fundamental
La traducción del código
Irreversibilidad de la traducción
Perturbaciones microscópicas
Incertidumbre operacional e incertidumbre esencial
La evolución: creación absoluta y no revelación
Desde su nacimiento, en las islas Jónicas, hace cerca de tres mil años, el
pensamiento occidental se ha repartido entre dos actitudes en apariencia
opuestas. Según una de esas filosofías, la realidad auténtica y última del
universo no puede residir más que en formas perfectamente inmutables,
invariantes por esencia. Según la otra, al contrario, es en el movimiento y la
evolución donde reside la única realidad del universo.
Platón y Heráclito
De Platón a Whitehead, y de Heráclito a Hegel y Marx, es evidente que estas
epistemologías metafísicas han estado siempre íntimamente asociadas a las ideas
morales y políticas de sus autores. Estos edificios ideológicos, presentados
como a
priori,
eran en realidad construcciones a
posteriori destinadas
a justificar una teoría ético-política preconcebida.[28]
El único a
priori,
para la ciencia, es el postulado de objetividad que le ahorra, o más bien le
prohíbe, tomar parte en este debate. La ciencia estudia la evolución, sea la del
universo o la de los sistemas que contiene, como el de la biosfera, comprendido
el hombre. Sabemos que todo fenómeno, todo acontecimiento, todo conocimiento,
implica interacciones, por sí mismas generadoras de modificaciones en los
componentes del sistema. Esta noción, sin embargo, no es en ningún modo
incompatible con la idea que existe de las entidades inmutables en la estructura
del universo. Más bien al contrario: la estrategia fundamental de la ciencia en
el análisis de los fenómenos es el descubrimiento de los invariantes. Toda ley
física, como además todo desarrollo matemático, especifica una relación de
invariancia; las proposiciones más fundamentales de la ciencia son postulados
universales de conservación. Es fácil ver, en todo ejemplo que se quiera
escoger, que es de hecho imposible analizar un fenómeno cualquiera en otros
términos que los de los invariantes conservados por este fenómeno. El ejemplo
más claro es quizá la formulación de las leyes de la cinética, que exigió la
invención de las ecuaciones diferenciales, es decir, de un medio para definir el
cambio en términos de lo que permanece sin cambiar.
Se puede ciertamente preguntar si todas las invariancias, conservaciones y
simetrías que constituyen la trama del discurso científico no son ficciones que
substituyen a la realidad para dar una imagen operacional, vacía por una parte
de substancia, pero que se vuelve accesible a una lógica fundada sobre un
principio de identidad puramente abstracto, quizá «convencional». Convención de
la que, sin embargo, la razón humana parece incapaz de abstenerse.
Menciono aquí este problema clásico para hacer notar que su estatuto ha sido
profundamente modificado por la revolución cuántica. El principio de identidad
no figura como postulado físico en la ciencia clásica. No es empleado más que
como operación lógica, sin que sea necesario suponer que corresponde a una
realidad substancial. La diferencia es absoluta por lo que respecta a la física
moderna, de la que uno de los postulados fundamentales es la identidad absoluta de
dos átomos que se encuentren en el mismo estado cuántico.[29] De
ello proviene, igualmente, el valor de representación absoluta, no perfectible,
otorgado a las simetrías atómicas y moleculares en teoría cuántica. Parece,
pues, que hoy no se pueda restringir el principio de identidad al estatuto de
simple regla para la conducta del espíritu: es preciso admitir que al menos a
escala cuántica expresa una realidad substancial.
Sea cual sea, existe y existirá en la ciencia un elemento platónico que no se
podría extirpar sin arruinarla. En la diversidad infinita de los fenómenos
singulares, la ciencia no puede buscar más que las invariantes.
Las invariantes anatómicas
Había una ambición «platónica» en la búsqueda sistemática de las invariantes
anatómicas a la que se consagraron los grandes naturalistas del siglo XIX, tras
Cuvier (y Goethe). Quizá los biólogos modernos no siempre reconocen el genio de
los hombres que, bajo la estupefaciente variedad de las morfologías y de los
modos de vida de los seres vivos, supieron descubrir si no una «forma» única, al
menos un número finito de planos anatómicos, cada uno de ellos invariante en el
seno del grupo que él caracteriza. Sin duda no es muy difícil ver que las focas
son mamíferos muy próximos a los carnívoros terrestres; pero lo es mucho más
discernir un mismo plano fundamental en la anatomía de los tunicados y de los
vertebrados, para agruparlos en la rama de los cordados. Y aún más difícil el
percibir las afinidades entre los cordados y los equinodermos. Sin embargo, no
hay duda alguna (la bioquímica lo confirma), que los erizos de mar son parientes
más próximos nuestros, que los miembros de ciertos grupos más evolucionados,
tales como los cefalópodos, por ejemplo.
Es gracias a estos inmensos trabajos, a la búsqueda de los planos fundamentales
de organización, que ha sido levantado el edificio de la zoología clásica y de
la paleontología, cuya estructura reclama y justifica a la vez la teoría de la
evolución. La diversidad de los tipos, sin embargo, permanecía, y era preciso
reconocer que numerosos planos de organizaciones macroscópicas, radicalmente
diferentes unos de otros, coexistían en la biosfera. Entre un alga azul, un
infusorio, un pulpo y el hombre, por ejemplo, ¿qué hay de común? El
descubrimiento de la célula y la teoría celular permitían entrever una nueva
unidad bajo esta diversidad. Sin embargo, fue preciso esperar los avances de la
bioquímica en el curso del segundo cuarto del siglo XX, principalmente, para que
se revelara enteramente la profunda y rigurosa unidad, a escala microscópica, de
todo el mundo viviente. Se sabe hoy en día que, desde la bacteria al hombre, la
maquinaria química es esencialmente la misma, tanto por sus estructuras como por
su funcionamiento.
Las invariantes químicas
-
En su estructura: todos los seres vivos, sin excepción, están constituidos
de las mismas dos clases principales de macromoléculas: proteínas y ácidos
nucleicos. Además, estas macromoléculas están formadas, en todos los seres
vivos, por el ensamblaje de los mismos radicales, en número finito: veinte
aminoácidos para las proteínas, cuatro tipos de nucleótidos para los ácidos
nucleicos.
-
Por su funcionamiento: las mismas reacciones, o más bien secuencias de
reacciones, son utilizadas en todos los organismos para las operaciones
químicas esenciales: movilización y puesta en reserva del potencial químico,
biosíntesis de los constituyentes celulares.
Ciertamente, en este tema central del metabolismo se encuentran numerosas
variantes, que corresponden a diversas adaptaciones funcionales. Casi siempre,
sin embargo, consisten en utilizaciones nuevas de secuencias metabólicas
universales, primeramente empleadas en otras funciones. Por ejemplo, la
excreción del nitrógeno se hace bajo formas diferentes en las aves y en los
mamíferos. Las aves excretan ácido úrico, los mamíferos urea. O sea que la vía
de síntesis del ácido úrico en las aves no es más que una pequeña modificación
de la secuencia de reacciones que, en todos los organismos, sintetiza los
nucleótidos llamados púricos (constituyentes universales de los ácidos
nucleicos). La síntesis de la urea, en los mamíferos, es obtenida gracias a una
modificación de una vía metabólica igualmente universal: la que culmina en la
síntesis de la arginina, aminoácido presente en todas las proteínas. Podríamos
multiplicar sin dificultad los ejemplos. Es a los biólogos de mi generación a
los que les ha sido revelada la casi identidad de la química celular en la
biosfera entera. Desde 1950 la certidumbre era plena, y cada publicación nueva
aportaba la confirmación. Las esperanzas de los «platónicos» más convencidos
estaban más que satisfechas.
Pero esta revelación, gradual, de la «forma» universal de la química celular,
parecía por otra parte hacer más agudo y más paradójico aún el problema de la
invariancia reproductiva. Si, químicamente, los constituyentes son los mismos, y
sintetizados por las mismas vías en todos los seres vivos, ¿cuál es la fuente de
su prodigiosa diversidad morfológica y fisiológica? Y más aún, ¿cómo cada
especie, utilizando los mismos materiales y las mismas transformaciones químicas
que todas las demás, mantiene, invariante a través de las generaciones, la norma
estructural que la caracteriza y la diferencia de cualquier otra?
Poseemos hoy la solución de este problema. Los constituyentes universales que
son por una parte los nucleótidos, y por otra los aminoácidos, son el
equivalente lógico de un alfabeto con el que estaría la estructura, o sea las
funciones asociativas específicas de las proteínas. Con este alfabeto puede por
lo tanto ser escrita toda la diversidad de las estructuras de las performances que
contiene la biosfera. Además, es la reproducción, ne
varietur,
en cada generación celular del texto escrito bajo forma de secuencia de
nucleótidos en el ADN, que asegura la invariancia de la especie.
El ADN como invariante fundamental
La invariante biológica fundamental es el ADN. Por ese motivo, la definición,
por Mendel, del gen como portador invariante de los rasgos hereditarios, su
identificación química por Avery (confirmada por Hershey) y la elucidación, por
Watson y Crick, de las bases estructurales de su invariancia replicativa,
constituyen sin ninguna duda los descubrimientos más fundamentales que hayan
sido hechos jamás en biología. A los que es preciso añadir la teoría de la
evolución selectiva que, además, no podía encontrar toda su significación y su
certidumbre más que gracias a estos descubrimientos.
La estructura del ADN, cómo esta estructura da cuenta de su capacidad de dictar
una copia exacta de la secuencia de nucleótidos de un segmento de ADN en una
secuencia de aminoácidos en una proteína; todos estos hechos y nociones han sido
amplia y excelentemente expuestos por los no especialistas. No los volveremos a
tratar aquí en detalle[30].
El siguiente esquema, que simboliza solamente la esencia de los dos procesos de replicación y
de traducción bastará
básicamente para la presente discusión;
 [31] [31]
El primer punto que conviene aclarar, es que el «secreto» de la replicación ne
varietur del
ADN reside en la complementariedad
estéreo-química del
complejo no
covalente que
constituyen las dos fibras asociadas en la molécula. Se ve, pues, que el
principio fundamental de estéreo-especificidad asociativa, que rinde cuenta de
las propiedades discriminativas de las proteínas, está igualmente en la base de
las propiedades replicativas del ADN. Pero en el ADN la estructura topológica
del complejo es mucho más simple que en los complejos de proteínas, y esto es lo
que permite funcionar a la mecánica de la replicación. En efecto, la estructura
estereoquímica de una de las dos fibras está enteramente definida por la
secuencia (la sucesión) de los radicales que la componen, en virtud del hecho de
que cada
uno de
los cuatro radicales no es individualmente apareable
(en razón de las restricciones estéticas), más que con uno
solo de
los otros tres. De ello resulta que:
-
la estructura estérica del complejo puede ser completamente representada
en dos dimensiones, de las que una, finita, contiene en cada punto un
par de nucleóticos mutuamente complementarios, mientras que la otra contiene
una secuencia potencialmente infinita de estos pares;
-
teniendo una (cualquiera) de las dos fibras, podrá ser reconstituida la
secuencia complementaria progresivamente por adiciones sucesivas de
nucleótidos, siendo «elegido» cada uno por su compañero estéricamente
predestinado. Es así como cada una de las dos fibras dicta la estructura de su
complementaria para reconstituir el complejo entero.
La estructura global de la molécula de ADN es la más simple y la más probable
que pueda adoptar una macromolécula constituida por la polimerización lineal de
radicales parecidos: la de una fibra helicoidal, definida por dos operaciones de
simetría, una traslación y una rotación. Se la puede considerar, pues, como un
cristal fibrilar, en razón de la regularidad de la estructura de conjunto. Pero
si se observa la estructura fina se verá que se trata de un cristal aperiódico,
ya que la secuencia de los pares de bases no es repetitiva. Es importante
subrayar que la secuencia es totalmente «libre», en el sentido que ninguna
restricción es impuesta por la estructura de conjunto, que puede acomodarse a
todas las secuencias posibles.
Como se acaba de ver, la formación de esta estructura es muy comparable a la de
un cristal. Cada elemento de secuencia en una de las dos fibras representa el
papel de un germen cristalino, que elige y orienta las moléculas que vienen
espontáneamente a asociarse, asegurando el crecimiento del cristal. Dos fibras
complementarias, artificialmente disociadas, reforman espontáneamente el
complejo específico, escogiendo cada una, casi sin errores, su compañera entre
millares o millones de secuencias.
Sin embargo, el crecimiento de cada fibra implica la formación de uniones covalentes que
asocian secuencialmente los nucleótidos entre sí. La formación de estas uniones
no puede tener lugar espontáneamente: le hace falta una fuente de potencial
químico y un catalizador. La fuente de potencial está representada por ciertas
uniones, presentes en los mismos nucleótidos, y que se rompen en el curso de la
reacción de condensación. Ésta viene catalizada por una enzima, la ADN-polimerasa.
Esta enzima es «indiferente» a la secuencia, especificada por la fibra
preexistente. Ha sido probado, además, que la condensación de mononucleótidos
activada por catalizadores no enzimáticos, es efectivamente dirigida por su
apareo espontáneo con un polinucleótido preexistente.[32][ Es
cierto, sin embargo, que si la enzima no especifica la secuencia, contribuye a
la precisión de la copia complementaria, es decir a la fidelidad del traspaso de
información. Fidelidad extrema, como lo prueba la experiencia pero que,
tratándose de un proceso microscópico, no puede ser absoluta. Volveremos pronto
sobre este punto capital.
La traducción del código
El mecanismo de la traducción de
la secuencia de nucleótidos en secuencia de aminoácidos es mucho más complicado,
en su principio mismo, que el de la replicación.
Este último proceso se explica en definitiva, como se acaba de ver, por
interacciones estéreo-específicas directas entre
una secuencia polinucleotídica que sirve de matriz y los nucleótidos que vienen
a asociarse. Igualmente en la traducción, son interacciones estéreo-específicas
no covalentes las que aseguran la transferencia de información, Pero estas
interacciones directrices comprenden varias etapas sucesivas, poniendo en juego
varios constituyentes que reconocen cada uno, exclusivamente, su compañero
funcional inmediato. Los constituyentes que intervienen al principio de esta
cadena de transporte de información ignoran totalmente «lo que sucede» en el
otro extremo. De suerte que, aunque es cierto que el código genético está
escrito en un lenguaje estéreo-químico, en el que cada letra está constituida
por una secuencia de tres nucleótidos (un triplete) en el ADN, específicos para
un aminoácido (entre veinte) en el polipéptido, no existe ninguna relación
estérica directa entre el triplete codificante y el aminoácido codificado.
Esto entraña una conclusión muy importante: que este código, universal en la
biosfera, parece químicamente arbitrario,
en el sentido de que la transferencia de información podría perfectamente tener
lugar según otra convención.[33] Se
conocen además mutaciones que, alterando la estructura de algunos componentes
del mecanismo de traducción, modifican la interpretación de algunos tripletes y
cometen, pues (en relación con la convención reinante), errores muy
perjudiciales al organismo.
El aspecto muy mecánico, e incluso «tecnológico», del proceso de traducción
merece ser subrayado. Las interacciones sucesivas de los diferentes componentes
que intervienen en cada etapa para desembocar en un polipéptido en vía de
ensamblaje, residuo a residuo, en la superficie de un constituyente (el
ribosoma) comparable a una máquina herramienta que hace avanzar muesca a muesca
una pieza que va a ser conformada, hacen pensar irresistiblemente en la cadena
de producción de una fábrica de mecánica.
En conjunto, en el organismo normal, esta mecánica microscópica de precisión
confiere al proceso de traducción una notable fidelidad. Sin duda hay errores,
pero tan poco probables que no se posee sobre su frecuencia normal media ninguna
estadística utilizable. Suponiendo al código sin ambigüedad (para la traducción
del ADN en proteínas), se deduce que la secuencia de los nucleótidos en un
segmento de ADN define enteramente la secuencia de los aminoácidos en el
polipéptido correspondiente. Como además, y ya se vio antes (capítulo 5), la
secuencia del polipéptido especifica enteramente (en condiciones iniciales
normales) la estructura doblada que adopta una vez constituido, la
«interpretación» estructural, o sea funcional, de la información genética es
univoca, rigurosa. Ningún aporte suplementario de información (que no sea
genético) es necesario, ni incluso parece posible; el mecanismo tal como se le
conoce no deja lugar para ello. Y en la medida en que todas las estructuras y performances de
los organismos son la resultante de las estructuras y actividades de las
proteínas que los componen, se debe considerar que el organismo entero
constituye la expresión epigenética última del mismo mensaje genético.
Irreversibilidad de la traducción
Es preciso añadir, finalmente, y este punto es de una gran importancia, que el
mecanismo de la traducción es estrictamente irreversible.
Ni se ha observado, ni es concebible, que la «información» sea jamás transferida
en el sentido inverso, es decir de proteína a ADN. Esta noción se apoya en un
conjunto de observaciones tan completas y tan seguras hoy en día, y sus
consecuencias en la teoría de la evolución, principalmente, son tan importantes,
que se la debe considerar como uno de los principios fundamentales de la moderna
biología. De ello se deduce, en efecto, que no hay mecanismo posible por
el que la estructura y las performances pudieran
ser modificadas y estas modificaciones transmitidas, aunque sólo fuera
parcialmente, a la descendencia, si no es como consecuencia de una alteración de
las instrucciones representadas por un segmento de secuencia del ADN. Mientras
que inversamente no existe ningún mecanismo concebible por el que una
instrucción o información cualquiera pudiera ser transferido al ADN.
El sistema entero, por consecuencia, es totalmente, intensamente conservador,
cerrado sobre sí mismo, y absolutamente incapaz de recibir cualquier instrucción
del mundo exterior. Como se ve, este sistema, por sus propiedades, por su
funcionamiento de relojería microscópica que establece entre ADN y proteína,
como también entre organismo y medio, relaciones de sentido único, desafía toda
descripción «dialéctica». Es profundamente cartesiano y no hegeliano: la célula
es una máquina.
Podría, pues, parecer que, por su misma estructura, este sistema deba oponerse a
todo cambio, a toda evolución. Nadie duda que no sea exactamente así, y aquí
tenemos la explicación[34] de
un hecho en verdad más paradójico que la misma evolución, a saber, la prodigiosa
estabilidad de ciertas especies, que han sabido reproducirse sin modificaciones
apreciables desde hace centenas de millones de años.
* * * *
La física sin embargo no enseña que (salvo en el cero absoluto, limite
inaccesible) toda entidad microscópica puede sufrir perturbaciones de orden
cuántico, cuya acumulación, en el seno de un sistema macroscópico, alterará la
estructura, de forma gradual pero infalible.
Los seres vivos, pese a la perfección conservadora de la maquinaria que asegura
la fidelidad de la traducción, no escapan a esta ley. El envejecimiento y la
muerte de los organismos pluricelulares se explican, al menos en parte, por la
acumulación de errores accidentales de traducción que, alterando principalmente
ciertos componentes responsables de la fidelidad de la traducción, acrecientan
la frecuencia de estos errores, que degradan poco a poco, inexorablemente, la
estructura de estos organismos.[35]
Perturbaciones microscópicas
El mecanismo de la replicación no podría tampoco, sin violar las leyes de la
física, escapar a toda perturbación, a todo accidente. Al menos, algunas de
estas perturbaciones entrañarán modificaciones más o menos discretas de ciertos
elementos de secuencia. Errores de transcripción que, en virtud de la fidelidad
ciega del mecanismo, serán, junto a otras perturbaciones, automáticamente re
transcritos. Serán fielmente traducidos en una alteración de la secuencia de los
aminoácidos en el polipéptido correspondiente al segmento de ADN en el que se
producirá la mutación.
Mas hasta que este polipéptido parcialmente nuevo se repliegue sobre sí mismo no
se revelará la «significación» funcional de la mutación.
Entre las modernas investigaciones en biología, algunas de las más hermosas por
su metodología, como también más profundamente significativas, constituyen lo
que se llama la genética molecular (Benzer, Yanofsky, Brenner y Crick). Estas
investigaciones han permitido, en particular, analizar los diferentes tipos de
alteraciones accidentales discretas que puede sufrir una secuencia de
polinucleótidos en la doble fibra del ADN. Se han identificado así diversas
mutaciones como debidas a:
-
la sustitución de un solo par de nucleótidos por otro;
-
la supresión o la adición de uno o varios pares de nucleótidos;
-
diversos tipos de «mástiques» que alteran el texto genético por inversión,
repetición, transposición y fusión de segmentos de secuencia más o menos
largos.[36]
Decimos que estas alteraciones son accidentales, que tienen lugar al azar. Y ya
que constituyen la
única fuente
posible de modificaciones del texto genético, único depositario,
a su vez, de las estructuras hereditarias del organismo, se deduce
necesariamente que sólo el
azar está en el origen de toda novedad, de toda creación en la biosfera. El puro
azar, el único azar, libertad absoluta pero ciega, en la raíz misma del
prodigioso edificio de la evolución: esta noción central de la biología moderna
no es ya hoy en día una hipótesis, entre otras posibles o al menos, concebibles.
Es la
sola concebible,
como única compatible con los hechos de observación y experiencia. Y nada
permite suponer (o esperar) que nuestras concepciones sobre este punto deberán o
incluso podrán ser revisadas.
Esta noción es, también, de todas las de todas las ciencias, la más destructiva
de todo antropocentrismo, la más inaceptable intuitivamente para los seres
intensamente teleonómicos que somos nosotros. Es pues la noción, o más bien el
espectro, que debe a toda costa exorcizar todas las ideologías vitalistas y
animistas. También es muy importante precisar en qué sentido exacto puede y debe
ser empleada la palabra azar, tratándose de las mutaciones como fuente de la
evolución. El contenido de la noción de azar no es simple, y la misma palabra se
emplea en situaciones muy diferentes. Lo mejor es tomar algunos ejemplos.
Incertidumbre operacional e incertidumbre esencial
Se emplea esta palabra, por ejemplo, a propósito del juego de dados, o de la
ruleta, y se utiliza el cálculo de probabilidades para prever el resultado de
una jugada. Pero estos juegos puramente mecánicos, y macroscópicos,
no son «de azar» más que en razón de la imposibilidad práctica de
gobernar con una precisión suficiente el lanzamiento del dado o el de la bola.
Es evidente que un mecanismo de lanzamiento de muy alta precisión es concebible,
y permitiría eliminar en gran parte la incertidumbre del resultado. Digamos que,
en la ruleta, la incertidumbre es puramente operacional, pero no esencial.
Ocurre igual, como se verá fácilmente, en la teoría de numerosos fenómenos donde
se emplea la noción de azar y el cálculo de probabilidades por razones puramente
metodológicas.
Pero, en otras situaciones, la noción de azar toma una significación esencial y
no ya simplemente operacional. Es el caso, por ejemplo, de lo que se pueden
llamar las «coincidencias absolutas», es decir, las que resultan de la
intersección de dos cadenas causales totalmente independientes una de otra.
Supongamos, por ejemplo, que el Dr. Dupont sea llamado urgentemente para visitar
a un nuevo enfermo, mientras que el plomero Dubois trabaja en la reparación
urgente de la techumbre de un inmueble vecino. Cuando el Dr. Dupont pasa por
debajo del alero del inmueble, el plomero suelta por inadvertencia su martillo,
cuya trayectoria (determinista) es interceptada por la del médico, que muere con
el cráneo roto. Decimos que no hubo suerte. ¿Qué otro término emplear para un
acontecimiento así, imprevisible por su misma naturaleza? El azar aquí debe
evidentemente ser considerado como esencial, inherente a la independencia total
de las dos series de acontecimientos cuyo encuentro produjo el accidente.
De modo que entre los acontecimientos que pueden provocar o permitir un error en
la replicación del
mensaje genético y sus consecuencias funcionales, hay igualmente independencia
total. El efecto funcional depende de la estructura, del papel actual de la
proteína modificada, de las interacciones que asegura, de las reacciones que
cataliza. Cosas todas que no tienen nada que ver con el acontecimiento
mutacional, como con sus causas inmediatas o lejanas, y, además, con la
naturaleza determinista o no de estas «causas».
Existe, en fin, a escala microscópica, una fuente de incertidumbre más radical
aún, enraizada en la estructura cuántica de la misma materia. Luego una mutación
es en sí un acontecimiento microscópico, cuántico, al que por consecuencia se
aplica el principio de incertidumbre. Acontecimiento pues esencialmente imprevisible
por su misma naturaleza.
Como se sabe, el principio de incertidumbre no ha sido jamás enteramente
aceptado por algunos de los más grandes físicos modernos, empezando por Einstein
que decía no poder admitir que «Dios juegue a los dados». Ciertas escuelas han
querido no ver más que una noción puramente operacional, pero no esencial. Todos
los esfuerzos hechos para sustituir por una estructura más «fina» la teoría
cuántica, con lo que la incertidumbre habría desaparecido, son sin embargo
liquidados y muy pocos físicos parecen dispuestos a creer hoy en día que este
principio podrá desaparecer de su disciplina.
Pase lo que pase es preciso subrayar que, aunque el principio de incertidumbre
deba un día ser abandonado, todo permanecerá de modo que entre el determinismo,
aunque sea total, de una mutación de secuencia en el ADN y sus efectos
funcionales a nivel de las interacciones de la proteína, no se podrá ver más que
una «coincidencia absoluta», en el sentido definido más arriba por la parábola
del plomero y del doctor. El acontecimiento se quedará pues en el dominio del
azar «esencial». A menos, claro, de volver al universo de Laplace, donde el azar
está excluido por definición y donde el doctor, siempre, debía morir bajo el
martillo del plomero.
La evolución: creación absoluta y no revelación
Bergson, acordémonos, veía en la evolución la expresión de una fuerza creadora, absoluta en
el sentido que no la suponía orientada a otro fin que la creación misma y por sí
misma. En esto difiere radicalmente de los animistas (trátese de Engels, de
Teilhard o de los positivistas optimistas como Spencer), que ven en la evolución
el majestuoso desarrollo de un programa inscrito en la misma trama del Universo.
Para ellos, en consecuencia, la evolución no es verdaderamente creación, sino
únicamente «revelación» de las intenciones, hasta entonces inexpresadas, de la
naturaleza. De aquí la tendencia a ver en el desarrollo embrionario una
emergencia del mismo orden que la emergencia evolutiva. Según la teoría moderna,
la noción de «revelación» se aplica al desarrollo epigenético, pero no, desde
luego, a la emergencia evolutiva que, gracias precisamente al hecho de que su
fuente está en lo imprevisible esencial, es creadora de novedad absoluta. Esta
convergencia aparente entre las vías de la metafísica bergsoniana y las de la
ciencia, ¿será también el efecto de una pura coincidencia? Quizá no: Bergson,
como artista y poeta que era, muy bien informado además de las ciencias
naturales de su tiempo, no podía dejar de ser sensible a la deslumbrante riqueza
de la biosfera, a la variedad prodigiosa de las formas y de los comportamientos
que en ella se despliegan, y que parecen testimoniar casi directamente, en
efecto, una prodigalidad creadora inagotable, libre de toda sujeción.
Pero allá donde Bergson veía la prueba más manifiesta de que el «principio de la
vida» es la misma evolución, la biología moderna reconoce, al contrario, que
todas las propiedades de los seres vivos reposan sobre un mecanismo fundamental
de conservación
molecular.
Para la teoría moderna, la
evolución no es de ningún modo una propiedad de los seres vivos,
ya que tiene su raíz en las imperfecciones
mismas del
mecanismo conservador que constituye su único privilegio. Es preciso, pues,
decir que la misma fuente de perturbaciones, de «ruido» que, en un sistema no
vivo, es decir no replicativo, aboliría poco a poco toda estructura, es el
origen de la evolución en la biosfera, y demuestra su total libertad creadora,
gracias a este conservatorio del azar, sordo al ruido tanto como a la música: la
estructura replicativa del ADN.
Capítulo 7
Evolución
Contenido:
El azar y la necesidad
Riqueza de la fuente de azar
«Paradoja» de la estabilidad de las especies
La irreversibilidad de la evolución y el segundo principio
Origen de los anticuerpos
El comportamiento como orientando las presiones de selección
El lenguaje y la evolución del hombre
La adquisición primaria del lenguaje
La adquisición del lenguaje programado en el desarrollo epigenético del
cerebro
Los acontecimientos elementales iniciales que abren la vía de la evolución a
esos sistemas intensamente conservadores que son los seres vivos, son
microscópicos, fortuitos y sin ninguna relación con los efectos que puedan
entrañar en el funcionamiento teleonómico.
El azar y la necesidad
Pero, una vez inscrito en la estructura del ADN, el accidente singular, y como
tal esencialmente imprevisible, va a ser mecánica y fielmente replicado y
traducido, es decir a la vez multiplicado y transpuesto a millones o a miles de
millones de ejemplares. Sacado del reino del puro azar, entra en el de la
necesidad, de las certidumbres más implacables. Porque es a escala macroscópica,
la del organismo, a la que opera la selección.
Muchos espíritus distinguidos, aún hoy, parecen no poder aceptar, ni incluso
comprender, que de una fuente de ruido la selección haya podido, ella sola,
sacar todas las músicas de la biosfera. La selección opera, en efecto, sobre los
productos del azar y no puede alimentarse de otra forma; pero opera en un
dominio de exigencias rigurosas donde el azar es desterrado. Es de estas
exigencias, y no del azar, de donde la evolución ha sacado sus orientaciones
generalmente ascendentes, sus conquistas sucesivas, el crecimiento ordenado del
que ella parece dar la imagen.
Algunos evolucionistas post-darwinianos han tenido, además, la tendencia de
propagar una idea empobrecida de la selección natural, una idea ingenuamente
feroz: la de la pura y simple «lucha por la vida», expresión, además, que no es
de Darwin, sino de Spencer. Los neo-darwinianos de principio de siglo han
propuesto, al contrario, una concepción mucho más rica y han mostrado, sobre la
base de teorías cuantitativas, que el factor decisivo de la selección no es la
«lucha por la vida» sino, en el seno de una especie, la tasa diferencial de
reproducción.
Los datos de la biología contemporánea permiten aclarar y precisar aún más la
noción de selección. Tenemos, particularmente, de la potencia, de la complejidad
y de la coherencia de la red cibernética intracelular (incluso en los organismos
más simples) una idea bastante clara, antes ignorada, que nos permite, mucho
mejor que antiguamente, comprender que toda «novedad», bajo la forma de una
alteración de la estructura de una proteína, podrá o no ser aceptada en función
de su compatibilidad con el conjunto de un sistema ya ligado por innumerables
sujeciones que deciden la ejecución del proyecto del organismo. Las únicas
mutaciones aceptables son, pues, las que, por lo menos, no reducen la coherencia
del aparato teleonómico, sino más bien lo confirman en la orientación ya
adoptada o, sin duda más raramente, lo enriquecen de nuevas posibilidades.
El aparato teleonómico, tal como funciona cuando se expresa por primera vez una
mutación, es el que define las condiciones
iniciales esenciales
de la admisión, temporal o definitiva, o del rechazo de la tentativa nacida del
azar. Es la performance teleonómica,
expresión global de las propiedades de la red de las interacciones constructivas
y reguladoras, la que es juzgada por la selección, y es de este hecho que la
evolución parece cumplir un «proyecto»: el de prolongar y amplificar un «sueño»
ancestral.
Riqueza de la fuente de azar
Gracias a la perfección conservadora del aparato replicativo, toda mutación,
considerada individualmente, es un acontecimiento muy raro. En las bacterias,
únicos organismos de los que se tienen numerosos y precisos datos en esta
materia, se puede admitir que la probabilidad, para un gen dado, de sufrir una
mutación que altere sensiblemente las propiedades funcionales de la proteína
correspondiente, es del orden de 10−6 a
10−8 por
generación celular. Pero en algunos mililitros de agua una población de varios
miles de millones de células puede desarrollarse. En una población así, se
tiene, pues, la certeza de que toda mutación dada está representada por 10, 100
o 1.000 ejemplares. Se puede estimar igualmente que el número total de mutantes
de todas las especies de esta población es del orden de 105 a
106.
A escala de la población, en consecuencia, la mutación no es en ningún modo un
fenómeno excepcional; es la regla. O sea que es en el seno de la población, pero
no en los individuos aislados, donde se ejerce la presión de selección. Las
poblaciones de organismos superiores, es cierto, no alcanzan las dimensiones que
las de las bacterias, pero:
-
El genoma de un organismo superior, por ejemplo un mamífero, contiene mil
veces más genes que el de una bacteria;
-
el número de generaciones celulares, o sea de probabilidades de
mutaciones, en la línea germinal de óvulo a óvulo o de espermatozoide a
espermatozoide es muy grande.
Es quizá lo que explica que la tasa de algunas mutaciones en el hombre parezca
relativamente elevada: del orden de 10−4 a
10−5 por
ejemplo, para un cierto número de mutaciones, que provocan enfermedades
genéticas fácilmente señalables. También es preciso hacer notar que las cifras
aquí dadas no tienen en cuenta las mutaciones individualmente no descubribles,
pero que, asociadas por recombinación sexual, podrían tener sensibles efectos.
Es probable que tales mutaciones hayan tenido más importancia en la evolución
que aquellas cuyos efectos individuales son más marcados.
En total, se puede estimar que, en la población humana actual (3×109)
se producen, en cada generación, de unos cien a mil millares de millones de
mutaciones. No doy esta cifra más que para dar una idea de las dimensiones del
inmenso depósito de variabilidad fortuita que constituye el genoma de una
especie, pese, una vez más, a las propiedades celosamente conservadoras del
mecanismo replicativo.
«Paradoja» de la estabilidad de las especies
Teniendo en cuenta las dimensiones de esta enorme lotería y la velocidad a la
que actúa la naturaleza, no es ya la evolución, sino al contrario la estabilidad
de las «formas» en la biosfera lo que podría parecer difícilmente explicable, si
no casi paradójico.
Se sabe que los planes de organización que corresponden a las principales
ramificaciones del reino animal estaban diferenciados desde el fin del Cámbrico,
o sea hace 500 millones de años. Se sabe también que incluso ciertas especies no
han evolucionado sensiblemente desde centenas de millones de años. La lingula,
por ejemplo, desde hace 450 millones de años; en cuanto a la ostra de hace 150
millones de años, debía tener la misma apariencia y el mismo sabor que las que
se sirven en los restaurantes.[37] En
fin, se puede estimar que la célula «moderna», caracterizada por su plan de
organización química invariante (empezando por la estructura del código genético
y el mecanismo complicado de la traducción), existe desde hace dos o tres miles
de millones de años, ya provista sin ninguna duda de potentes redes cibernéticas
moleculares asegurando su coherencia funcional.
La extraordinaria estabilidad de algunas especies, los miles de millones de años
que cubre la evolución, la invariancia del «plan» químico fundamental de la
célula no pueden evidentemente explicarse más que por la extrema coherencia del
sistema teleonómico que, en la evolución, ha jugado pues el papel a la vez de
guía y de freno, y no ha retenido, amplificado, integrado más que una ínfima
fracción de las probabilidades que le ofrecía, en número astronómico, la ruleta
de la naturaleza.
El sistema replicativo, por su parte, lejos de poder eliminar las perturbaciones
microscópicas de las que es inevitablemente el objeto, no sabe al contrario más
que registrarlas y ofrecerlas, casi siempre vanamente, al filtro teleonómico
cuyas performances son
juzgadas, en último caso, por la selección.
* * * *
Una mutación simple, puntual, como la sustitución de una letra del código por
otra, en el ADN, es reversible. La teoría lo prevé, y la experiencia lo prueba.
Pero toda evolución sensible, como la diferenciación de dos especies, incluso
muy vecinas, resulta de un gran número de mutaciones independientes,
sucesivamente acumuladas en la especie original, y después, siempre al azar,
recombinadas gracias al «flujo genético» promovido por la sexualidad. Un
fenómeno así, en razón del número de los acontecimientos independientes del que
resulta, es estáticamente irreversible.
La irreversibilidad de la evolución y el segundo principio
La evolución en la biosfera es pues un proceso necesariamente irreversible, que
define una dirección en el tiempo;
dirección que es la
misma que
la que impone la ley de crecimiento de la entropía, es decir, el segundo
principio de la termodinámica. Se trata de mucho más que de una comparación. El
segundo principio está fundado sobre consideraciones estadísticas idénticas a
las que establecen la irreversibilidad de la evolución. De hecho, es
legítimo considerar la irreversibilidad de la evolución como una expresión del
segundo principio en la biosfera.
El segundo principio, no formulando más que una predicción estadística, no
excluye, desde luego, que un sistema macroscópico cualquiera no pueda, en un
movimiento de muy débil amplitud y con una duración muy corta, remontar la
cuesta de la entropía, es decir, en cierto modo remontar el tiempo. En los seres
vivos, son precisamente estos únicos y fugitivos movimientos los que, captados y
reproducidos por el mecanismo replicativo, han sido retenidos por la selección.
En este sentido la evolución selectiva, fundada sobre la elección de los raros y
preciosos incidentes que también contiene, entre infinidad de otros, el inmenso
depósito del azar microscópico, constituye una especie de máquina de remontar el
tiempo.
No es sorprendente, sino al contrario muy natural, que los resultados obtenidos
por este mecanismo de remontar el tiempo: la tendencia general ascendente de la
evolución, el perfeccionamiento y el enriquecimiento del aparato teleonómico,
hayan parecido milagrosos a unos, paradójicos a otros y que la teoría moderna «darwiniano-molecular»
de la evolución sea hoy todavía considerada con suspicacia por algunos
pensadores, filósofos o incluso biólogos.
Origen de los anticuerpos
Esto resulta, al menos por una parte, de la extrema dificultad que existe en
concebir la inagotable riqueza de la fuente de azar donde bebe la selección.
Hay, sin embargo, un ejemplo destacable en el sistema de defensa del organismo
por los anticuerpos.
Los anticuerpos son proteínas dotadas de la propiedad de reconocer por
asociación estéreo-específica substancias extrañas al organismo y que lo han
invadido, bacterias o virus, por ejemplo. Pero, como sabemos, el anticuerpo que
reconoce electivamente una substancia dada, por ejemplo un «motivo estérico»
particular a una cierta especie bacteriana, no aparece en el organismo (para
habitarlo durante un cierto tiempo) más que después que éste haya hecho, al
menos una vez, la «experiencia» (por la vacunación, espontánea o artificial). Se
ha demostrado, además, que el organismo es capaz de formar anticuerpos adaptados
prácticamente a no importa qué motivo estérico, natural o sintético. Las
potencialidades, en este aspecto, parecen prácticamente infinitas.
Se ha supuesto, pues, durante largo tiempo, que la fuente de información para la
síntesis de la estructura asociativa específica del anticuerpo era el mismo
antígeno. Pero está establecido hoy que la estructura del anticuerpo no debe
nada al antígeno: en el seno del organismo células especializadas, producidas en
gran número, poseen la propiedad —única— de «jugar a la ruleta» en una parte,
bien definida, de los segmentos genéticos que determinan la estructura de los
anticuerpos. El funcionamiento exacto de esta ruleta genética especializada y
ultrarrápida no está aún enteramente elucidado: es verosímil, sin embargo, que
intervengan tanto recombinaciones como mutaciones, unas y otras en todo caso
produciéndose al azar, en la ignorancia total de la estructura del antígeno.
Éste, por el contrario, juega el papel de selector, favoreciendo
diferencialmente la multiplicación de las células capaces de producir un
anticuerpo que pueda reconocerle.
Es muy notable encontrar, en la base de uno de los fenómenos de adaptación
molecular más exquisitamente precisos que se conoce, una fuente al azar. Pero
está claro (a
posteriori) que
sólo una fuente así puede ser lo bastante rica para ofrecer al organismo medios
de defensa en cierto modo «acimutales».
El comportamiento como orientando las presiones de selección
Otra dificultad para la teoría selectiva proviene de haber sido demasiado a
menudo comprendida o presentada como dependiente de las solas condiciones del medio
exterior como
agentes de la selección. Ésta es una concepción completamente errónea.
Porque las presiones de selección que ejercen sobre los organismos las
condiciones externas, no son en ningún caso independientes de las performances teleonómicas
características de la especie. Organismos diferentes viviendo en el mismo
«nicho» ecológico, tienen con las condiciones externas (comprendidos los demás
organismos), interacciones muy diferentes y especificas. Son estas interacciones
específicas, en parte «escogidas» por el mismo organismo, las que determinan la
naturaleza y la orientación de la presión de selección que él sufre. Digamos que
las «condiciones iniciales» de selección que encuentra una mutación nueva,
comprenden a la vez, y de forma indisoluble, el medio exterior y el conjunto de
las estructuras y performances del
aparato teleonómico.
Es evidente que el papel de las performances teleonómicas
en la orientación de la selección es cada vez mayor a medida que se eleva el
nivel de organización, es decir de autonomía del
organismo respecto al medio. Y esto, al extremo que se pueda considerar sin duda
este papel como decisivo en los organismos superiores, cuya supervivencia y
reproducción dependen ante todo de su comportamiento.
Es, además, evidente que la elección inicial de uno y otro tipo de
comportamiento podrá a menudo tener una influencia de gran duración, no sólo
para la especie donde será manifestada por primera vez, sino para toda su
descendencia, aunque constituya un grupo entero. Como se sabe, las grandes
articulaciones de la evolución han sido debidas a la invasión de nuevos espacios
ecológicos. Si los vertebrados tetrápodos han aparecido y han podido dar la
maravillosa expansión que representan los anfibios, los reptiles, las aves y los
mamíferos, es porque en el origen, un pez primitivo «eligió» el ir a explorar la
tierra donde no podía sin embargo desplazarse más que saltando dificultosamente.
Él creó así, como consecuencia de una modificación de comportamiento, la presión
de selección que debía desarrollar los poderosos miembros de los tetrápodos.
Entre los descendientes de este explorador audaz, este Magallanes de la
evolución, algunos pueden correr a más de 70 km/h, otros trepan a los árboles
con una sorprendente agilidad, otros en fin han conquistado el aire, cumpliendo,
prolongando, amplificando de modo prodigioso el «sueño» del pez ancestral.
El hecho de que, en la evolución de algunos grupos, se observe una tendencia
general, sostenida durante millones de años, al desarrollo aparentemente
orientado de ciertos órganos, atestigua que la elección inicial de un cierto
tipo de comportamiento (ante la agresión de un predador por ejemplo) compromete
a la especie a permanecer en la vía de un perfeccionamiento continuo de las
estructuras y performances que
son el soporte de este comportamiento. Debido a que los antecesores del caballo
habían escogido vivir en la llanura y huir al acercarse un predador (antes que
intentar defenderse o esconderse), la especie moderna, tras una larga evolución
que comprende múltiples estados de reducción, anda hoy sobre el extremo de un
solo dedo.
Se sabe que ciertos comportamientos muy precisos y complejos, como las
ceremonias prenupciales de las aves, están estrechamente emparejados a ciertas
características morfológicas particularmente llamativas. Es cierto que la
evolución de este comportamiento y la del carácter anatómico sobre la que reposa
van a la par, una atrayendo y reforzando a la otra bajo la presión de la
selección sexual. Desde que ella comienza a desarrollarse en una especie, todo
adorno asociado al éxito del acoplamiento no hace más que reforzar, confirmar en
suma, la presión de selección inicial, y en consecuencia favorecer todo
perfeccionamiento de este mismo adorno. Es pues legítimo decir que es el
instinto sexual, en suma, el deseo,
el que ha creado las condiciones de selección de algunos magníficos plumajes.[38]
Lamarck pensaba que la tensión de los esfuerzos desplegados por un animal para
«triunfar en la vida» actuaba en algún modo sobre su patrimonio hereditario para
incorporarse y modelar directamente su descendencia. El inmenso cuello de la
jirafa expresaba en suma la voluntad constante que habían tenido sus antepasados
de llegar a las ramas más altas de los árboles. Hipótesis hoy en día
inaceptable, desde luego, pero que muestra que la pura selección, operando sobre
los elementos del comportamiento, desemboca en el resultado que Lamarck quería
expresar el estrecho emparejamiento de las adaptaciones anatómicas y de las performances específicas.
El lenguaje y la evolución del hombre
Es en estos términos en los que es preciso considerar el problema de las
presiones de selección que han orientado la evolución del hombre. Problema de un
interés excepcional, independientemente incluso del hecho de que se trate de
nosotros, y que al discernir mejor en su evolución las raíces de nuestro ser,
pudiera llegarse a comprenderlo mejor en su actual naturaleza. Porque un
observador imparcial, un marciano por ejemplo, debería sin ninguna duda
reconocer que el desarrollo de la performance específica
del hombre, el lenguaje simbólico, acontecimiento único en la biosfera, abre el
camino a otra evolución,
creadora de un nuevo reino, el de la cultura, de las ideas, del conocimiento.
Acontecimiento único: los lingüistas modernos han insistido en el hecho de que
el lenguaje simbólico del hombre es absolutamente irreductible a los medios de
comunicación tan diversos (auditivos, táctiles, visuales u otros) empleados por
los animales. Actitud sin ninguna duda justificada. Pero de aquí a afirmar que
la discontinuidad en la evolución ha sido absoluta, que el lenguaje humano desde
el origen no
debía estrictamente nada, por ejemplo, a un sistema de llamadas y avisos
variados como los que intercambian los grandes simios, me parece un paso difícil
de franquear, en todo caso una hipótesis inútil.
El cerebro de los animales es, sin duda alguna, capaz, no sólo de registrar
informaciones, sino también de asociarlas y transformarlas, y de restituir el
resultado de estas operaciones bajo forma de una performance individual;
pero no, y éste no es el punto esencial, bajo una forma que permita comunicar a
otro individuo una asociación o transformación original, personal. Esto es,
contrariamente a lo que permite el lenguaje humano, lo que hace que se le pueda
considerar por definición como nacido el día en que combinaciones creadoras,
asociaciones nuevas,
realizadas en un individuo, pudieron, transmitidas a otros, no perecer con él.
No se conocen lenguas primitivas: en todas las razas de nuestra única especie
moderna, el instrumento simbólico ha llegado sensiblemente al mismo nivel de
complejidad y poder de comunicación. Según Chomsky, además, la estructura
profunda, la «forma» de todas las lenguas humanas es la misma. Las
extraordinarias performances que
la lengua a la vez representa y autoriza, están evidentemente asociadas al
desarrollo considerable del sistema nervioso central en el Homo
sapiens;
desarrollo que constituye además su rasgo anatómico más distintivo.
Se puede hoy afirmar que la evolución del hombre, desde sus más antiguos
antecesores conocidos, se ha centrado ante todo en el desarrollo progresivo de
la caja craneana, o sea del cerebro.
Ha sido necesario para ello una presión de selección orientada, continuada, y
sostenida desde más de dos millones de años. Presión de selección considerable,
porque esta duración es relativamente corta, y específica porque
no se ve nada parecido en ninguna otra raza: la capacidad craneana de los simios
antropomorfos modernos no es más grande casi que la de sus antecesores de hace
algunos millones de años.
Es imposible no suponer que entre la evolución privilegiada del sistema nervioso
central del hombre y la de la performance única
que lo caracteriza, no hubiese habido un emparejamiento muy estrecho, que hizo
del lenguaje no sólo el producto, sino una de las condiciones iniciales de esta
evolución.
La hipótesis que me parece más verosímil es que, aparecida muy prontamente en
nuestra raza, la comunicación simbólica más rudimentaria, por las posibilidades
radicalmente nuevas que ofrecía, constituyó una de estas «elecciones» iniciales
que comprometen el porvenir de la especie creando una presión de selección
nueva; esta selección debía favorecer el desarrollo de la performance lingüística
y por consecuencia la del órgano que la sirve, el cerebro. Hay, creo, a favor de
esta hipótesis, argumentos bastante poderosos.
Los hominianos auténticos más antiguos que se conocen hoy (los Australopitecos,
que Leroi-Gourhan, con razón prefiere llamar Australántropos) poseían ya, lo que
los definía además, las características que distinguen al hombre de sus más
próximos parientes, los póngidos (es decir los simios antropoides). Los
Australántropos habían adoptado la posición erecta, asociada no sólo a una
especialización del pie, sino a numerosas modificaciones del esqueleto y de la
musculatura, principalmente de la columna vertebral y de la posición del cráneo
en relación a aquélla. Se ha insistido a menudo sobre la importancia que ha
debido tener, en la evolución del hombre, la liberalización de las servidumbres
de la marcha a cuatro patas practicada por todos los antropoides, a excepción
sin embargo del gibón. Nadie duda que esta invención, muy antigua (anterior a
los Australántropos), haya tenido una extrema importancia: sólo ella podía
permitir a nuestros antepasados convertirse en cazadores capaces, sin dejar de
andar o de correr, de utilizar sus miembros anteriores.
La capacidad craneana de estos hominianos primitivos era sin embargo apenas
superior a la de un chimpancé y ligeramente inferior a la de un gorila. El peso
del cerebro no es seguramente proporcional a sus performances.
Nadie duda, sin embargo, que le impone un límite, y que el Homo
sapiens no
hubiese podido emerger sino gracias al desarrollo de su caja craneana.
Sea lo que sea, parece establecido que si el cerebro del Zijántropo no pesaba
más que el del gorila, era, sin embargo capaz de performances desconocidas
para los póngidos: El Zijántropo, en efecto, tenía una industria, tan primitiva
a decir verdad que sus «útiles» no son reconocibles como artefactos más que por
la repetición de las mismas estructuras, muy rudimentarias, y por su
agrupamiento alrededor de ciertos esqueletos fósiles. Los grandes simios emplean
«útiles» naturales, piedras o ramas de árboles, cuando se presenta la ocasión,
pero no producen nada comparable a artefactos confeccionados según una norma reconocible.
Así el Zijántropo debe ser considerado como un Homo
faber muy
primitivo. Luego, parece muy verosímil, que entre el desarrollo del lenguaje y
el de una industria testimoniando una actividad proyectiva y disciplinada, deba
haber una correlación muy estrecha.[39] Parece
pues razonable suponer que los Australántropos poseían un instrumento de
comunicación simbólica a la medida de su industria rudimentaria. Además, si es
cierto como piensa Dart,[40] que
los Australántropos cazaban con éxito, como otros animales, bestias poderosas y
peligrosas como el rinoceronte, el hipopótamo y la pantera, sería preciso que
esto fuera una performance convenida
de antemano por un grupo de cazadores. Proyecto cuya formulación previa habría
exigido el empleo de un lenguaje.
A esta hipótesis parecía oponerse el débil desarrollo en volumen del cerebro de
los Australántropos. Pero recientes experiencias con un joven chimpancé parecen
mostrar que, si los simios son incapaces de aprender el lenguaje articulado,
pueden asimilar y utilizar algunos elementos del lenguaje simbólico de los
sordomudos.[41] Podemos
entonces suponer que la adquisición del poder de simbolización articulada ha
podido depender de modificaciones neuromotrices, no necesariamente muy
complejas, en un animal que era, en este estado, más inteligente que un
chimpancé actual.
Pero es evidente que una vez franqueado este paso, el uso de un lenguaje, por
primitivo que sea, no puede dejar de acrecentar en proporciones considerables el
valor de supervivencia de la inteligencia, y de crear en favor del desarrollo
del cerebro una precisión de selección poderosa y orientada, tal como ninguna
especie afásica podía jamás esperar. En el momento que existe un sistema de
comunicación simbólica, los individuos, o más bien los grupos mejor dotados para
su empleo, adquieren sobre los otros una ventaja incomparablemente más grande
que la que les habría conferido una igual superioridad de inteligencia a
individuos de una especie desprovista de lenguaje. Vemos también que la presión
de selección debida al uso de un lenguaje debe favorecer especialmente la
evolución del sistema nervioso central en el sentido de una inteligencia de un
cierto tipo: el que es más apto para explotar esta performance particular,
específica, dueña de inmensos poderes.
La adquisición primaria del lenguaje
Esta hipótesis sólo seria atrayente y razonable si no hubiese además adquirido
singular importancia al haber sido reclamada por algunos datos relativos al
actual lenguaje. El estudio de la adquisición del lenguaje por el niño sugiere
de modo irresistible[42] que
si este proceso nos parece milagroso se debe a que es, por naturaleza,
profundamente diferente del aprendizaje regular de un sistema de reglas
formales. El niño no aprende regla alguna, ni siquiera intenta imitar el
lenguaje de los adultos. Podría decirse que en cada estadio de su desarrollo
toma del lenguaje aquello que le conviene. En el estadio primerísimo (hacia el
decimoctavo mes), el niño puede poseer un stock de diez palabras, que siempre
usa aisladamente, sin asociarlas jamás, ni por imitación. Más tarde asociará las
palabras de dos en dos, o de tres en tres, etc., de acuerdo con una síntesis que
tampoco es simple repetición o imitación del lenguaje adulto. Este proceso
parece que es universal y que su cronología es idéntica para todas las lenguas.
La facilidad con que, en dos o tres años (después del primero), este juego del
niño con la lengua le da sobre ella un perfecto dominio parece siempre increíble
al observador adulto.
Por eso es difícil no ver el reflejo de un proceso embriológico, epigenético, en
el curso del cual se desarrollan las estructuras neurales subyacentes a las performances lingüísticas.
Esta hipótesis se ve confirmada por las observaciones relativas a las afasias de
origen traumático. Cuando se dan en el niño, estas afasias retroceden tanto más
rápidamente y más completamente cuanto más joven sea el pequeño. Estas lesiones
son, al contrario, irreversibles cuando ocurren en la pubertad o más tarde. Todo
un conjunto de observaciones, además de las aducidas, confirman que existe una
edad crítica para la adquisición espontánea del lenguaje. Cada cual sabe
perfectamente que aprender una segunda lengua en la edad adulta exige un
esfuerzo voluntario sistemático y sostenido. El estatuto de la lengua aprendida
en estas condiciones permanece siempre prácticamente inferior al de la lengua
natal, aprendida espontáneamente.
La adquisición del lenguaje programado en el desarrollo
epigenético del cerebro
La idea de que la adquisición primaria del lenguaje está vinculada a un proceso
de desarrollo epigenético se ve confirmada por los datos anatómicos. En efecto,
se sabe que la maduración del cerebro prosigue después del nacimiento para
acabar en la pubertad. Este desarrollo parece consistir esencialmente en un
enriquecimiento considerable de las intercomunicaciones de las neuronas
corticales. Este proceso, muy rápido durante los dos primeros años, se retrasa
después. No continúa (visiblemente) más allá de la pubertad y cubre, por
consiguiente, el «período crítico» durante el que la adquisición primaria es
posible.[43]
De esto a pensar que si, en el niño, la adquisición del lenguaje parece tan
milagrosamente espontánea es porque se inscribe en la misma trama de desarrollo
epigenético, una
de cuyas funciones es acogerle,
entonces sólo hay un paso que, por mi parte, no dudo en dar. Para intentar ser
un poco más preciso: de este crecimiento postnatal del córtex depende, sin duda
alguna, el desarrollo de la función cognitiva. Es la adquisición del lenguaje en
el curso mismo de esta epigénesis lo que permitiría asociarlo a la función
cognitiva y esto de un modo tan íntimo que es muy difícil para nosotros
disociar, por la introspección, la performance lingüística
del conocimiento que explícita.
Se admite en general que el lenguaje no constituye más que una
«superestructura», a lo que se llega, claro está, por la extrema diversidad de
las lenguas humanas, productos de la segunda evolución, la de la cultura. Sin
embargo, la amplitud y el refinamiento en el Homo
sapiens de
las funciones cognitivas no encuentran, evidentemente, su razón de ser más que y
por el lenguaje. Privadas de este instrumento, son, en su mayor parte,
inutilizables, paralizadas. En este sentido la capacidad lingüística ya no puede
ser considerada como una superestructura. Es preciso admitir que entre las
funciones cognitivas y el lenguaje simbólico, que ellas reclaman y por las que
se explicitan, hay en el hombre moderno una estrecha simbiosis, que no puede ser
producto más que de una larga evolución común.
Se sabe que, según Chomsky y su escuela, bajo la extrema diversidad de las
lenguas humanas, el análisis lingüístico en profundidad revela una «forma» común
a todas ellas. Esta forma debe pues, según Chomsky, ser considerada como innata y
característica de la especie. Esta concepción ha escandalizado a algunos
filósofos o antropólogos que ven en ella un retomo a la metafísica cartesiana. A
condición de aceptar el contenido biológico implícito, esta condición no me
choca en absoluto. Ella me parece, al contrario, natural, a partir del momento
en que se admite que la evolución de las estructuras corticales del hombre no ha
podido dejar de ser influenciada, en una parte importante, por una capacidad
lingüística adquirida de modo muy temprano en el estadio más rudimentario. Lo
que es lo mismo que admitir que el lenguaje articulado, desde su aparición en el
linaje humano, no ha permitido solamente la evolución de la cultura, sino ha
contribuido de modo decisivo a la evolución física del
hombre. Si ha sucedido así, la capacidad lingüística que se revela en el curso
del desarrollo epigenético del cerebro forma parte actualmente de la «naturaleza
humana» definida en el seno del genoma en un lenguaje radicalmente diferente del
código genético. ¿Milagro? Ciertamente, puesto que en última instancia se trata
de un producto del azar. Pero el día en que el Zijántropo, o cualquiera de sus
compañeros, usó por vez primera un símbolo articulado para representar una
categoría, aumentó por este hecho en proporciones inmensas la probabilidad de
que un día emergiera un cerebro capaz de concebir la Teoría darwiniana de la
Evolución.
Capítulo 8
Las fronteras
Contenido:
Las fronteras actuales del conocimiento biológico
El problema de los orígenes
El enigma del origen del código
La otra frontera: el sistema nervioso central
Funciones del sistema nervioso central
El análisis de las impresiones sensoriales
Innatismo y empirismo
La función de simulación
La ilusión dualista y la presencia del espíritu
Cuando se piensa en el inmenso camino recorrido por la evolución a lo largo de
tres mil millones de años, en la prodigiosa riqueza de las estructuras que ha
creado, en la milagrosa eficacia de las performances de
los seres vivos, de la bacteria al hombre, se puede empezar a dudar que todo
ello sea producto de una enorme lotería, sacando números al azar, entre los
cuales una selección ciega ha designado los escasos ganadores.
Las fronteras actuales del conocimiento biológico
Al repasar en sus detalles las pruebas hoy acumuladas y ver que esta concepción
es la única compatible con los hechos (principalmente con los mecanismos
moleculares de la replicación, de la mutación y de la traducción), se halla la
certidumbre, pero no sin embargo una comprensión inmediata, sintética e
intuitiva, de la evolución en su conjunto. El milagro está «explicado»: nos
parece aún milagroso. Como escribió Mauriac: «Lo que dice este profesor es mucho
más increíble aún que lo que nosotros, pobres cristianos, creemos.»
Es cierto, como también lo es que no llegamos a formamos una imagen mental
satisfactoria de algunas abstracciones de la física moderna. Pero sabemos
también que tales dificultades no pueden tomarse como argumento contra una
teoría basada en las certidumbres de la experiencia y de la lógica. Para la
física, microscópica o cosmológica, vemos la causa de la incomprensión
intuitiva: la escala de los fenómenos considerados trasciende las categorías de
nuestra experiencia inmediata. Sólo la abstracción puede suplir esta
incapacidad, sin curarla. Para la biología la dificultad es de otro orden. Las
interacciones elementales sobre las que reposa toda la trama son relativamente
fáciles de comprender gracias a su carácter mecanístico. Es la fenomenal
complejidad de los sistemas vivientes lo que desafía toda representación
intuitiva global. En biología como en física, no hay, en estas dificultades
subjetivas, argumento contra la teoría.
Se puede decir hoy en día que los mecanismos elementales de la evolución están
no sólo comprendidos en principio, sino identificados con precisión. La solución
encontrada es tanto más satisfactoria cuanto que se trata de los mecanismos que
aseguran la estabilidad de las especies: invariancia replicativa del ADN,
coherencia teleonómica de los organismos.
La evolución sigue siendo en biología la noción central, destinada a
enriquecerse y precisarse durante mucho tiempo aún. Sin embargo, en lo esencial,
el problema está resuelto y la evolución no figura ya en las fronteras del
conocimiento.
Por mi parte, veo estas fronteras situadas en los dos extremos de la evolución:
el origen de los primeros sistemas vivientes por un lado, y por otro el
funcionamiento del sistema más intensamente teleonómico que jamás haya surgido;
me refiero al sistema nervioso central del hombre. En el presente capítulo,
querría intentar delimitar estas dos fronteras de lo desconocido.
El problema de los orígenes
Se podría pensar que el descubrimiento de los mecanismos universales sobre los
que reposan las propiedades esenciales de los seres vivos ha facilitado la
solución del problema de los orígenes. De hecho, estos descubrimientos,
renovando casi enteramente la cuestión, planteada hoy en términos mucho más
precisos, la han revelado más difícil todavía de lo que antes parecía.
Se pueden a
priori definir
tres etapas en el proceso que ha podido conducir a la aparición de los primeros
organismos:
-
la formación en la tierra de los constituyentes químicos esenciales de los
seres vivos, nucleótidos y aminoácidos;
-
la formación, a partir de estos materiales, de las primeras macromoléculas
de replicación;
-
la evolución que, alrededor de estas «estructuras replicativas», ha
construido un aparato teleonómico, hasta llegar a la célula primitiva.
Los problemas que plantea la interpretación de cada una de estas etapas son
diferentes. La primera, llamada a menudo la fase «prebiótica», es ampliamente
accesible, no sólo en la teoría, sino en la experiencia. Si la incertidumbre
continúa, y continuará sin duda, en lo tocante a las vías que ha seguido de
hecho la evolución química prebiótica, el cuadro de conjunto, por otra parte,
parece bastante claro. Las condiciones de la atmósfera y de la corteza
terrestre, hace cuatro mil millones de años, eran favorables a la acumulación de
ciertos compuestos simples del carbono, tales como el metano. Había también agua
y amoníaco. Y, de estos compuestos simples y en presencia de catalizadores no
biológicos, se obtienen bastante fácilmente numerosos cuerpos más complejos,
entre los que figuran aminoácidos y precursores de los nucleótidos (bases
nitrogenadas, azúcares). El hecho notable es que, en ciertas condiciones, en que
la reunión parece muy plausible, el rendimiento de estas síntesis en cuerpos
idénticos o análogos a los constituyentes de la célula moderna, es muy elevado.
Se puede considerar pues como probado que,
en un momento dado, sobre la Tierra, algunas extensiones de agua podrían contener
en solución concentraciones elevadas de los constituyentes esenciales de las dos
clases de macromoléculas biológicas, ácidos nucleicos y proteínas. En esta «sopa
prebiótica» diversas macromoléculas podían formarse por polimerización de sus
precursores, aminoácidos y nucleótidos. En efecto, se han obtenido en
laboratorio, en condiciones «plausibles», polipéptidos y polinucleótidos
parecidos por su estructura general a las macromoléculas «modernas».
Hasta aquí, en consecuencia, no hay grandes dificultades. Pero la primera etapa
decisiva no está franqueada: la formación de macromoléculas capaces, en las
condiciones de la sopa primitiva, de promover su propia replicación sin el
auxilio de ningún aparato teleonómico. Esta dificultad no parece invencible. Se
ha mostrado que una secuencia polinucleotídica puede efectivamente guiar, por
emparejamiento espontáneo, la formación de elementos de secuencia
complementaria. Está claro que un mecanismo así sería muy ineficaz y sujeto a
innumerables errores. Pero, desde el momento en que entran en juego, los tres
procesos fundamentales de la evolución: replicación, mutación, selección,
comienzan a operar y dan una ventaja considerable a las macromoléculas más
aptas, por su estructura secuencial, para replicarse espontáneamente.[44]
La tercera etapa es, por hipótesis, la emergencia gradual de los sistemas
teleonómicos que, alrededor de la estructura replicativa, deben construir un organismo,
una célula primitiva. Es aquí cuando se llega verdaderamente a la «barrera del
sonido», porque no tenemos la más mínima idea de cómo podía ser la estructura de
una célula primitiva. El sistema viviente más simple que conocemos, la célula
bacteriana, pequeña maquinaria de una complejidad y una eficacia extremas,
alcanzó seguramente su presente estado de perfección hace unos mil millones de
años. El plan de conjunto de la química de esta célula es el mismo que el de
todos los demás seres vivientes. Ella emplea el mismo código genético y la misma
mecánica de traducción que, por ejemplo, las células humanas.
Así, las células más simples que nos es posible estudiar, no tienen nada de
«primitivo». Son el producto de una selección que ha podido, a través de
quinientos o mil millares de millones de generaciones, acumular un aparejo
teleonómico tan poderoso que los vestigios de las estructuras verdaderamente
primitivas son indiscernibles. Reconstruir, sin fósiles, semejante evolución es
imposible. De todos modos quería intentar sugerir una hipótesis plausible en
cuanto a la vía seguida por esta evolución, sobre todo en su punto de arranque.
El desarrollo del sistema metabólico, que debió, a medida que se empobrecía el
caldo primitivo, «aprender» a movilizar el potencial químico y a sintetizar los
constituyentes celulares, plantea ingentes problemas. Igual sucede para la
emergencia de la membrana de permeabilidad selectiva sin la que no puede haber
célula viable. Pero el mayor problema es el origen del código genético y del
mecanismo de su traducción. De hecho, no es de un «problema» de lo que debería
hablarse, sino más bien de un verdadero enigma.
El enigma del origen del código
El código no tiene sentido si no es traducido. La máquina traductora de la
célula moderna comporta al menos cincuenta constituyentes macromoleculares que
son ellos mismos codificados en el ADN: el código no puede ser traducido más que
por productos de traducción.
Es la expresión moderna de omne
vivum ex ovo.
¿Cuándo y cómo se cerró este anillo? Es demasiado difícil de imaginar. Pero el
hecho de que el código sea hoy descifrado y conocido por ser universal permite
al menos plantear el problema en términos precisos; simplificando un poco bajo
el aspecto de la siguiente alternativa:
-
la estructura del código se explica por razones químicas, o más
exactamente estereoquímicas; si un cierto código ha sido «escogido» para
representar un cierto aminoácido, es porque existe entre ellos una cierta
afinidad estereoquímica;
-
la estructura del código es químicamente arbitraria; el código, tal como
nosotros lo conocemos, resulta de una serie de elecciones al azar que poco a
poco lo han enriquecido.
La primera hipótesis parece con mucho la más seductora. En primer lugar porque
explicaría la universalidad del código. A continuación porque permitiría
imaginar un mecanismo primitivo de traducción en el que el alineamiento
secuencial de los aminoácidos para formar un polipéptido sería debido a una
interacción directa entre los aminoácidos y la estructura replicativa. En fin, y
principalmente, porque esta hipótesis, si fuera verdadera, seria en principio
verificable. Pero de las numerosas tentativas de verificación que han sido
hechas, el balance, por el momento, debe ser considerado como negativo.[45]
Quizá la última palabra sobre este asunto aún no haya sido pronunciada.
Esperando una confirmación que parece improbable, pasamos a la segunda
hipótesis, desagradable por razones metodológicas, lo que no significa en ningún
modo que sea inexacta. Desagradable por varias razones. No explica la
universalidad del código. Es preciso entonces admitir que, entre numerosas
tentativas de elaboración, una sola ha sobrevivido. Lo que, en sí, es muy
verosímil además, pero que no propone ningún modelo de traducción primitiva. La
especulación debe entonces suplirlo. Y no faltan algunas muy ingeniosas: el
campo está libre, demasiado libre. El enigma sigue, y encubre también la
respuesta a una pregunta de profundo interés. La vida ha aparecido sobre la
tierra: ¿cuál era antes del acontecimiento la probabilidad de que apareciera? No
queda excluida, al contrario, por la estructura actual de la biosfera, la
hipótesis de que el acontecimiento decisivo no se haya producido más que una
sola vez. Lo que significaría que su probabilidad a priori es casi nula.
Esta idea repugna a la mayoría de los hombres de ciencia. De un acontecimiento
único la ciencia no puede decir ni hacer nada. No puede «discurrir», formando
una clase, más que sobre acontecimientos cuya probabilidad a
priori,
por débil que sea, es finita. Además, por la universalidad misma de sus
estructuras, empezando por el código, la biosfera aparece como el producto de un
acontecimiento único. Es posible, desde luego, que este carácter singular se
deba a la eliminación, por la selección, de muchas otras tentativas o variantes.
Mas nada impone esta interpretación.
La probabilidad a
priori de
que se produzca un acontecimiento particular entre todos los acontecimientos
posibles en el universo, está próxima a cero. No obstante el universo existe; es
preciso que se produzcan acontecimientos, cuya probabilidad (antes del
acontecimiento) sea ínfima. No tenemos, en la hora actual, el derecho de
afirmar, ni el de negar, que la vida haya aparecido una
sola vez sobre
la Tierra, y que, por consecuencia, antes de que existiera, sus posibilidades de
ser fuesen casi nulas.
Esta idea no resulta desagradable sólo a los biólogos como hombres de ciencia.
Ella choca con nuestra tendencia humana de creer que toda cosa real en el
universo actual era necesaria, y desde siempre. Nos es preciso estar siempre en
guardia contra el sentimiento tan poderoso del destino. La ciencia moderna
ignora toda inmanencia. El destino se escribe a medida que se cumple, no antes.
El nuestro no lo estaba antes de que emergiera la especie humana, única en la
biosfera en la utilización de un sentido lógico de comunicación simbólica. Otro
acontecimiento único que debería, por eso mismo, prevenimos contra todo
antropocentrismo. Si fue único, como quizá lo fue la aparición de la misma vida,
sus posibilidades, antes de aparecer, eran casi nulas. El Universo no estaba
preñado de la vida, ni la biosfera del hombre. Nuestro número salió en el juego
de Montecarlo. ¿Qué hay de extraño en que, igual que quien acaba de ganar mil
millones, sintamos la rareza de nuestra condición?
La otra frontera: el sistema nervioso central
El lógico podría advertir al biólogo que sus esfuerzos para «comprender» el
entero funcionamiento del cerebro humano están condenados al fracaso porque
ningún sistema lógico sabe describir integralmente su propia estructura. Esta
advertencia estaría fuera de lugar, tan lejos se está aún de esta frontera
absoluta del conocimiento. De todas maneras esta objeción lógica no se aplica al
análisis por el hombre del sistema nervioso central de un animal. Sistema que se
puede suponer menos complejo y menos potente que el nuestro. Incluso en este
caso, sin embargo, existe una dificultad importante: la experiencia consciente
de un animal nos resulta impenetrable y sin duda siempre lo seguirá siendo. ¿Se
puede afirmar que una descripción exhaustiva del funcionamiento del cerebro de
una rana, por ejemplo, sería posible, en principio, mientras que aquel dato
permaneciera inaccesible? Se puede poner en duda. De modo que la exploración del
cerebro humano, pese a las barreras opuestas a la experimentación, permanecerá
siempre irreemplazable, por la posibilidad que ofrece de comparar los datos
objetivos y subjetivos relativos a una experiencia.
Pase lo que pase, la estructura y el funcionamiento del cerebro pueden y deben
ser explotados simultáneamente en todos los niveles accesibles con la esperanza
de que estas investigaciones, muy diferentes tanto por sus métodos como por su
objeto inmediato, convergerán un día. Por el momento ellas casi no convergen más
que por las dificultades que comportan.
Entre las más difíciles e importantes, están los problemas que plantea el
desarrollo epigenético de una estructura tan compleja como el sistema nervioso
central. En el hombre, comprende de 1012 a
1013 neuronas
interconectadas por medio de unas 1014 a
1015 sinapsis,
que asocian en algunos casos células nerviosas alejadas. He mencionado ya el
enigma que plantea la realización de interacciones morfo-genéticas a distancia y
no insistiré aquí sobre ello. Al menos tales problemas pueden ser claramente
planteados gracias, indudablemente, a ciertas notables experiencias.[46]
No se puede comprender el funcionamiento del sistema nervioso central a menos de
conocer el del elemento lógico primario que constituye la sinapsis. De todos los
niveles de análisis es el más accesible a la experiencia, y ciertas técnicas
refinadas han aportado una considerable masa de documentos. Se está lejos aún,
sin embargo, de una interpretación de la transmisión sináptica en términos de
interacciones moleculares. Problema esencial sin embargo, ya que es aquí sin
duda donde reside el último secreto de la memoria. Se ha propuesto desde hace
largo tiempo que éste sea registrado en forma de una alteración más o menos
irreversible de las interacciones moleculares responsables de la transmisión del
influjo nervioso a nivel de un conjunto de sinapsis. Esta teoría tiene mucha
verosimilitud, pero no pruebas directas.[47]
Pese a esta profunda ignorancia referente a los mecanismos primarios del sistema
nervioso central, la electrofisiología moderna ha aportado resultados
profundamente significantes sobre el análisis y la integración de las señales
nerviosas, principalmente en ciertas vías sensoriales.
En primer lugar sobre las propiedades de la neurona como integradora de las
señales que puede recibir (por intermedio de las sinapsis) de otras numerosas
células. El análisis ha probado que la neurona es estrechamente comparable, por
sus performances,
a los componentes integrados de una calculadora electrónica. Es capaz como éstos
de efectuar, por ejemplo, todas las operaciones lógicas del álgebra
proposicional. Pero además puede adicionar o sustraer diferentes señales
teniendo en cuenta su coincidencia en el tiempo, así como modificar la
frecuencia de
las señales que emite en función de la
amplitud de
las que recibe. De hecho, parece que ningún componente unitario, actualmente
utilizado por las modernas calculadoras, sea capaz de performances tan
variadas y finamente moduladas. Sin embargo la analogía entre las máquinas
cibernéticas y el sistema nervioso central es impresionante, y la comparación
fructuosa. Pero es preciso ver que se limita todavía a los niveles inferiores de
integración: primeros grados del análisis sensorial por ejemplo. Las funciones
superiores del córtex, en las que el lenguaje es la expresión, parecen aún
escaparse totalmente. Nos podríamos preguntar si no habrá más que una diferencia
«cuantitativa» (grado de complejidad), o quizá se trate de una diferencia
«cualitativa». Esta pregunta para mí no tiene sentido. Nada permite suponer que
las interacciones elementales sean de diferente naturaleza a diferentes niveles
de integración. Si hay un caso donde sea aplicable la primera ley de la
dialéctica, con seguridad es en éste.
Funciones del sistema nervioso central
El mismo refinamiento de las funciones cognitivas en el hombre, y la propagación
de las aplicaciones que de ellas hace, enmascaran las funciones primordiales que
llenan el cerebro de la serie animal (comprendido el hombre). Quizá se pueda
enumerar y definir estas funciones primordiales de la manera siguiente:
-
asegurar el pedido y la coordinación central de la actividad neuromotriz
en función, principalmente, de las correspondencias sensoriales;
-
contener, en forma de circuitos genéticamente determinados, programas de
acción más o menos complejos; liberarlos en función de estímulos particulares;
-
analizar, filtrar e integrar las correspondencias sensoriales para
construir una representación del mundo exterior adaptada a las performances específicas
del animal;
-
registrar los acontecimientos que (teniendo en cuenta la gama de performances específicas)
son significativos, agruparlos en clases, según sus analogías; asociar estas
clases según las relaciones (de coincidencia o de sucesión) de los
acontecimientos que las constituyen; enriquecer, refinar y diversificar los
programas innatos incluyendo estas experiencias;
-
imaginar, es decir, representar y simular acontecimientos
exteriores, o programas de acción del mismo animal.
Las funciones definidas por los tres primeros apartados son realizadas por el
sistema nervioso central de animales que no se califica generalmente de
superiores: artrópodos, por ejemplo. Los ejemplos más espectaculares que se
conocen de programas de acción innatos muy complejos se encuentran en los
insectos. Es dudoso que las funciones resumidas en el párrafo 4 desempeñen un
papel importante en estos animales.[48] Por
el contrario, contribuyen de manera muy importante al comportamiento de los
invertebrados superiores, como el pulpo[49] y
todos los vertebrados.
En cuanto a las funciones del párrafo 5, que se podrían llamar «proyectivas»,
son sin duda el privilegio de los vertebrados superiores. Pero, aquí, la barrera
de la conciencia se interpone, y puede ser que no sepamos reconocer los signos
exteriores de esta actividad (el sueño, por ejemplo) más que en nuestros
próximos parientes, sin que otras especies estén sin embargo privadas.
Las funciones 4 y 5 son cognitivas, mientras que las de los párrafos 1, 2 y 3
son únicamente coordinadoras y representativas. Sólo las funciones 5 pueden ser
creadoras de experiencia
subjetiva.
El análisis de las impresiones sensoriales
Según la proposición del párrafo 3, el análisis por el sistema nervioso central
de las impresiones sensoriales aporta una representación empobrecida y orientada
del mundo exterior. Una especie de resumen donde no figura con claridad lo que
particularmente interesa al animal en función de su comportamiento específico
(es, en suma, un resumen «crítico», tomando la palabra en una acepción
complementaria del sentido kantiano). La experiencia demuestra abundantemente
que así es realmente. Por ejemplo, el analizador situado detrás del ojo de una
rana le permite ver una mosca (es decir, un punto negro) en movimiento, pero no
en reposo.[50] De
suerte que la rana atrapará a la mosca sólo en vuelo. Es preciso insistir sobre
el hecho, probado por el análisis electro fisiológico, de que eso no es el
resultado de un comportamiento que haría desdeñar por la rana un punto negro
inmóvil, como no representativo, con certeza, de un alimento. La imagen del
punto inmóvil se imprime en la retina, pero no es transmitida;
el sistema no es excitado más que por un objeto en movimiento.
Ciertas experiencias con el gato[51] sugieren
una interpretación del hecho misterioso de que un campo reflejando a la vez
todos los colores del espectro sea visto como una playa blanca,
mientras que el blanco es subjetivamente interpretado como ausencia de todo
color. Los experimentadores han mostrado que, debido a unas inhibiciones
cruzadas entre ciertas neuronas, que responden respectivamente a las diversas
longitudes de onda, éstas no mandan señales cuando la retina está expuesta
uniformemente a la gama entera de las longitudes de onda visibles. Goethe, y no
Newton, tenía, en un sentido subjetivo, razón. Error eminentemente perdonable a
un poeta.
Que los animales sean capaces de clasificar objetos o relaciones entre objetos
según categorías abstractas, principalmente geométricas, no puede tampoco
dudarse: un pulpo o una rata pueden aprender la noción de triángulo, de círculo
o de cuadrado y reconocer sin error estas figuras por sus propiedades
geométricas, independientemente de la dimensión, de la orientación o del color
que pueda tener el objeto real que se les presente.
El estudio de los circuitos, que analizan las figuras presentadas en el campo de
visión del gato, demuestra que estas hazañas geométricas son debidas a la
estructura misma de los circuitos que filtran y recomponen, en definitiva, sus
propias restricciones a la imagen, de la que extraen ciertos elementos simples.
Algunas células nerviosas, por ejemplo, no responden más que a la figura de una
línea recta inclinada en sentido inverso. Las «nociones» de la geometría
elemental no están pues tan representadas en el objeto como por el analizador
sensorial, que lo percibe y recompone a partir de sus elementos más simples.[52][
Innatismo y empirismo
Estos modernos descubrimientos dan pues la razón, en un sentido nuevo, a
Descartes y a Kant, y la niegan al empirismo radical que sin embargo no ha
cesado casi de reinar en la ciencia desde hace doscientos años, sospechando de
toda hipótesis que suponía la «innatividad» de los cuadros del conocimiento. Aún
en nuestros días ciertos etólogos parecen ligados a la idea de que los elementos
del comportamiento, en el animal, son o bien innatos o bien aprendidos,
excluyendo cada modelo al otro. Esta concepción es enteramente errónea como
Lorenz ha demostrado enérgicamente.[53] Cuando
el comportamiento implica elementos adquiridos por la experiencia, lo son según
un programa que
es innato, es decir genéticamente determinado. La estructura del programa
convoca y guía el aprendizaje que se inscribirá pues en una cierta «forma»
preestablecida, definida en el patrimonio genético de la especie. Es sin duda
así cómo es preciso interpretar el proceso de aprendizaje primario del lenguaje
en el niño (cf. cap. 7). No hay ninguna razón para suponer que no sea igual para
las categorías fundamentales del conocimiento en el hombre, y quizá también para
el individuo y la sociedad. Tales problemas son en principio accesibles a la
experiencia. Los etólogos tratan casos parecidos todos los días. Experiencias
crueles, que es inimaginable practicar en el hombre, en el niño concretamente.
De modo que, por respeto a sí mismo, el hombre no puede más que prohibirse
explorar ciertas estructuras constitutivas de su ser.
* * * *
La larga controversia sobre la innatividad cartesiana de las «ideas», negada por
los empiristas, recuerda la que ha dividido a los biólogos con motivo de la
distinción entre fenotipo y genotipo. Distinción fundamental, indispensable a la
misma definición del patrimonio hereditario para los genetistas que la habían
introducido, pero muy sospechosa a los ojos de muchos biólogos no genetistas que
no veían en ella más que un artificio destinado a salvar el postulado de
invariancia del gen. Se encuentra aquí, una vez más, la oposición entre los que
no quieren conocer más que el objeto actual, concreto, en su entera presencia, y
los que buscan discernir la representación enmascarada de una forma ideal. No
hay más que dos clases de sabios, decía Alain: los que aman las ideas y los que
las aborrecen. Estas dos actitudes de espíritu se oponen todavía en la ciencia;
y son, una y otra, por su confrontación, necesarias a sus progresos. No se puede
más que lamentar, para los despreciadores de ideas, que este progreso, al que
contribuyen, invariablemente les perjudique.
En un sentido muy importante, los grandes empiristas del siglo XVIII no se
equivocaban sin embargo. Es perfectamente verdadero que todo, en los seres
vivos, viene de la experiencia, comprendida la innatividad genética, sea ésta la
del comportamiento estereotípico de las abejas o la de los cuadros innatos del
conocimiento humano. Pero no de la experiencia actual, renovada por cada
individuo, en cada generación: de la acumulada por la ascendencia entera de la
especie en el curso de la evolución. Sólo esta experiencia extraída al azar,
sólo estas tentativas innumerables, corregidas por la selección, podían, como de
todo otro órgano, hacer del sistema nervioso central un sistema adaptado a su
función particular. Para el cerebro: dar del mundo sensible una representación
adecuada a las performances de
la especie, suministrar el cuadro que permita clasificar eficazmente los datos
en sí mismos inutilizables de la experiencia inmediata e incluso, en el hombre,
simular subjetivamente la experiencia para anticipar los resultados y preparar
la acción.
La función de simulación
Son el poderoso desarrollo y el uso intensivo de la función de simulación los
que me parecen caracterizar las propiedades únicas del cerebro humano. Esto al
nivel más profundo de las funciones cognitivas, sobre el que reposa el lenguaje
y que sin duda no explícita sino en parte. Esta función sin embargo no es
exclusivamente humana. El perro, que manifiesta su alegría viendo a su amo
prepararse para el paseo, imagina evidentemente, es decir simula por
anticipación, los descubrimientos que hará, las aventuras que le esperan, los
espantos deliciosos que experimentará, sin peligro, gracias a la tranquilizante
presencia de su protector. Más tarde, simulará todo ello de nuevo, confusamente
mezclado, en sueños. En el animal, como también en el niño pequeño, la
simulación subjetiva no parece más que parcialmente disociada de la actividad
neuromotriz. Su ejercicio se traduce por el juego. Pero, en el hombre, la
simulación subjetiva deviene la función superior por excelencia, la función
creadora. Es ella la que es reflejada por la simbólica del lenguaje que la
explícita transponiendo y resumiendo sus operaciones. De ahí, el hecho,
subrayado por Chomsky, de que el lenguaje, incluso en sus más humildes empleos,
es casi siempre innovador: traduce una experiencia subjetiva, una simulación
particular, siempre nueva. Es en esto también en lo que el lenguaje humano
difiere radicalmente de la comunicación animal. Ésta se reduce a llamadas y
avisos que corresponden a un cierto número de situaciones concretas
estereotipadas. El animal más inteligente, capaz sin duda de simulaciones
subjetivas bastante precisas, no dispone de ningún medio de «liberar su
conciencia», si no es indicando groseramente en qué sentido anda
su imaginación. El hombre sabe narrar sus experiencias subjetivas: la
experiencia nueva, el encuentro creador no perece ya con aquél en que ella habrá
sido, por primera vez, simulado.
Todos los hombres de ciencia han debido, pienso yo, darse cuenta de que su
reflexión, a nivel profundo, es verbal: es una experiencia
imaginaria,
simulada con la ayuda de formas, de fuerzas, de interacciones que no componen
apenas una «imagen» en el sentido visual del término. Yo mismo me he
sorprendido, no teniendo, a fuerza de atención centrada en la experiencia
imaginaria, nada más en el campo de la conciencia, al identificarme a una
molécula de proteína. Sin embargo no es en este momento cuando aparece la
significación de la experiencia simulada, sino solamente una vez explicitada
simbólicamente. No creo en efecto que haga falta considerar las imágenes no
visuales sobre las que opera la simulación como símbolos, sino más bien, si me
atrevo a decirlo, como la «realidad» subjetiva y abstracta, directamente
ofrecida a la experiencia imaginaria.
Sea cual sea, en el uso corriente, el proceso de simulación está enteramente
enmascarado por la palabra que le sigue casi inmediatamente y parece confundirse
con el mismo pensamiento. Pero se sabe que numerosas observaciones objetivas
prueban que en el hombre las funciones cognitivas, incluso complejas, no están
inmediatamente ligadas a la palabra (o a cualquier otro medio de expresión
simbólica). Se pueden citar principalmente los estudios hechos sobre diversos
tipos de afasias. Quizá las experiencias más impresionantes son las, recientes,
de Sperry,[54] sobre
sujetos cuyos dos hemisferios cerebrales habían sido separados por sección
quirúrgica del corpus
callosum.
El ojo derecho y la mano derecha, en estos sujetos, no comunican información más
que al hemisferio izquierdo, y recíprocamente. Así, un objeto visto por el ojo
izquierdo, o palpado por la mano izquierda, es reconocido, sin que el sujeto
pueda nombrarlo. O en ciertos tests difíciles, donde se trataba de emparejar la
forma (tridimensional) de un objeto, tenido en una de las dos manos, al
desarrollo plano de
esta forma, representado en una pantalla, el hemisferio derecho (afásico) se
mostraba, con mucho, superior al hemisferio «dominante» (izquierdo), y más
rápido en la discriminación. Resulta tentador especular sobre la posibilidad de
que una parte importante, quizá la más «profunda» de la simulación objetiva,
esté asegurada por el hemisferio derecho.
* * * *
Si es legítimo considerar que el pensamiento reposa sobre un proceso de
simulación subjetiva, es preciso admitir que el alto desarrollo de esta facultad
en el hombre es resultado de una evolución en el curso de la cual es en la
acción concreta, preparada por la experiencia imaginaria, donde la eficacia de
este proceso, su valor de supervivencia, ha sido probado por la selección. Es
pues por su capacidad de representación adecuada y de previsión exacta,
confirmada por la experiencia concreta, por la que el poder de simulación del
sistema nervioso central, en nuestros antepasados, ha sido empujado hasta el
estado alcanzado en el Homo sapiens. El simulador subjetivo no tenía derecho a
equivocarse cuando se trataba de organizar una cacería de panteras con las armas
que disponía el Australántropo, el Pitecántropo, o incluso el Homo sapiens de
Cromagnon. Por eso es por lo que el instrumento lógico innato, heredado de
nuestros antepasados, no nos engaña y nos permite «comprender» los
acontecimientos del universo, es decir, describirnos en lenguaje simbólico y
preverlo, con tal de que los elementos de información necesarios sean
suministrados al simulador.
Instrumento de anticipación, enriqueciéndose sin cesar de los resultados de sus
propias experiencias, el simulador es el instrumento del descubrimiento y de la
creación. Es el análisis de la lógica de su funcionamiento subjetivo el que ha
permitido formular las reglas de la lógica objetiva y crear nuevos instrumentos
simbólicos, como las matemáticas. Grandes espíritus (Einstein) a menudo se
maravillan, con razón, del hecho de que los entes matemáticos creados por el
hombre puedan representar tan fielmente a la naturaleza sin deberle ellos nada a
la experiencia. Nada, es cierto, a la experiencia individual y concreta, pero
todo a las virtudes del simulador forjado por la experiencia innumerable y cruel
de nuestros humildes antepasados. Confrontando sistemáticamente la lógica y la
experiencia, según el método científico, es de hecho toda la experiencia de
nuestros antepasados la que confrontamos con la experiencia actual.
* * * *
Si podemos adivinar la existencia de este maravilloso instrumento, si sabemos
traducir, por el lenguaje, el resultado de sus operaciones, no tenemos ninguna
idea de su funcionamiento, de su estructura. La experimentación fisiológica es,
bajo este aspecto, importante todavía. La introspección, con todos sus peligros,
nos ha dicho pese a todo un poco más. Queda el análisis del lenguaje que sin
embargo no revela el proceso de simulación más que a través de transformaciones
desconocidas y no aclara, sin duda, todas sus operaciones.
La ilusión dualista y la presencia del espíritu
He aquí la frontera, casi tan infranqueable todavía para nosotros como lo era
para Descartes. En tanto no es franqueada, el dualismo conserva en suma su
verdad operacional. La noción de cerebro y la de espíritu no se confunden menos
ahora que en el siglo XVII. El análisis objetivo nos obliga a ver una ilusión en
el dualismo aparente del ser. Ilusión sin embargo tan íntimamente ligada al
mismo ser que sería sumamente vano esperar disiparla en la aprehensión inmediata
de la subjetividad, o aprender a vivir afectivamente, moralmente, sin ella. Y
además, ¿qué falta hace? ¿Quién podría dudar de la presencia del espíritu?
Renunciar a la ilusión que ve en el alma una «sustancia» inmaterial, no es negar
su existencia, sino al contrario comenzar a reconocer la complejidad, la
riqueza, la insondable profundidad de la herencia genética y cultural, como de
la experiencia personal, consciente o no, que en conjunto constituyen el ser que
somos, único e irrecusable testigo de sí mismo.
Capítulo 9
El reino y las tinieblas
Contenido:
Presiones de selección en la evolución del hombre
Peligros de degradación genética en las sociedades modernas
La selección de la ideas
La exigencia de explicación
Las ontogenias míticas y metafísicas
La ruptura de la «antigua alianza» animista y el mal del alma moderna
Los valores y el conocimiento
La ética del conocimiento
La ética del conocimiento y el ideal socialista
Presiones de selección en la evolución del hombre
Como ya dijimos, el día en que el Australántropo, o alguno de sus congéneres,
llegó a comunicar, no sólo una experiencia concreta y actual, sino el contenido
de una experiencia subjetiva, de una «simulación» personal, nació un nuevo
reino: el de las ideas. Una nueva evolución, la de la cultura, se hacía posible.
La evolución física del hombre debía proseguirse aún durante mucho tiempo,
estrechamente asociada en adelante a la del lenguaje, sufriendo profundamente su
influencia que trastornaba las condiciones de la selección.
El hombre moderno es el producto de esta simbiosis evolutiva. Él es
incomprensible, indescifrable, en cualquier otra hipótesis. Todo ser vivo es también un
fósil. Lleva en sí, y hasta en la estructura microscópica de sus proteínas, las
huellas, si no los estigmas, de su ascendencia. Esto es más cierto en el hombre
que en cualquier otra especie animal, en razón de la dualidad, física e «ideal»,
de la evolución de la que él es el heredero.
Se puede pensar que, durante centenas de milenios, la evolución ideal ha
precedido muy de cerca a la evolución física que la constreñía por el débil
desarrollo de un córtex capaz solamente de anticipar acontecimientos
directamente ligados a la inmediata supervivencia. De ahí, la intensa presión de
selección que debía empujar al desarrollo del poder de simulación y del lenguaje
que explícita las operaciones. Por ello también, la rapidez, sorprendente, de
esta evolución que testimonian los cráneos fósiles.
Pero a medida que proseguía esta evolución conjunta, su componente ideal no
podía más que independizarse respecto a los apremios que iniciaban poco a poco
el desarrollo del sistema nervioso central. Debido a esta evolución, el hombre
extendía su dominio sobre el universo subhumano y sufría menos los peligros que
le acechaban. La presión de selección que había guiado la primera fase de la
evolución podía entonces aflojarse y, en todo caso, tomar otro carácter.
Dominando en adelante sus cercanías, el hombre no tenía ya frente a sí más
adversario serio que él mismo. La lucha intraespecífica directa, la lucha a
muerte, se convertía desde entonces en uno de los principales factores de
selección en la especie humana. Fenómeno extremadamente raro en la evolución de
los animales. En el presente, la guerra intraespecífica, entre razas o grupos
distintos, es desconocida en las especies animales. En los grandes mamíferos,
incluso en el combate singular, frecuente entre machos, es extremadamente rara
la muerte del vencido. Todos los especialistas están de acuerdo en pensar que la
lucha directa, la struggle
for life de
Spencer, no ha jugado más que un pequeño papel en la evolución de las especies.
No sucede igual en el hombre. A partir al menos de un cierto grado de desarrollo
y de expansión de la especie, la guerra tribal o racial ha jugado evidentemente
un papel importante como factor de evolución. Es muy posible que la brutal
desaparición del hombre de Neanderthal fuera el resultado de un genocidio
cometido por nuestro antepasado Homo
sapiens.
Éste no debía ser el último: se conocen bastantes genocidios históricos.
¿En qué sentido esta presión de selección debía empujar la evolución humana?
Desde luego pudo favorecer la expansión de las razas mejor dotadas de
inteligencia, de imaginación, de voluntad, de ambición. Pero también debió
favorecer la cohesión de la banda, la agresividad del grupo más aún que el valor
solitario, y el respeto de las leyes de la tribu más que la iniciativa
individual.
Acepto todas las críticas que se quieran hacer a este esquema simplista. No
pretendo dividir la evolución humana en dos fases distintas. No he intentado más
que enumerar las principales presiones de selección que, ciertamente, han jugado
un papel muy importante en la evolución no sólo cultural, sino física del
hombre. El punto importante es que, durante centenares de miles de años, la
evolución cultural no podía dejar de influenciar la evolución física; en el
hombre, más aún que en cualquier otro animal, e incluso en razón de su autonomía
infinitamente superior, es el comportamiento el
que orienta la
presión de selección, Y a partir del momento en que el comportamiento cesa de
ser principalmente automático para hacerse cultural, los mismos rasgos
culturales ejercen su presión sobre la evolución del genoma.
Esto, sin embargo, hasta el momento en que la rapidez creciente de la evolución
cultural hace que ésta se disocie completamente de la del genoma.
Peligros de degradación genética en las sociedades modernas
Es evidente que, en el seno de las sociedades modernas, la disociación es total.
La selección ha sido suprimida. Al menos ya no tiene nada de «natural» en el
sentido darwiniano del término. En nuestras sociedades, y en la medida en que
rige todavía una selección, ella no favorece la «supervivencia del más apto», es
decir, en términos más modernos, la supervivencia genética del «más apto», por
una expansión más grande de su descendencia. La inteligencia, la ambición, el
coraje, la imaginación, son siempre factores de éxito en las sociedades
modernas. Pero de éxito personal,
y no genético,
que es el único que cuenta para la evolución. Como todos sabemos, las
estadísticas revelan una correlación negativa entre el cociente de inteligencia
(o el nivel de cultura) de los matrimonios y el número medio de hijos. Estas
mismas estadísticas demuestran por el contrario que existe, para el cociente de
inteligencia, una fuerte correlación positiva entre esposos. Situación
peligrosa, que corre el riesgo de atraer poco a poco hacia una elite,
que tendería en valor relativo a restringirse, el más elevado potencial
genético.
Todavía hay más: en una época reciente, incluso en las sociedades relativamente
«avanzadas», la eliminación de los menos aptos, física y también
intelectualmente, era automática y cruel. La mayoría no alcanzaba la pubertad.
Hoy, muchos de estos enfermizos genéticos sobreviven lo bastante como para
reproducirse. Gracias a los progresos del conocimiento y de la ética social, el
mecanismo que defendía a la especie contra la degradación, inevitable al
abolirse la selección natural, ya no funciona más que para las tareas muy
graves.
A estos peligros, a menudo señalados, se han opuesto a veces los remedios
alcanzados por los recientes progresos de la genética molecular. Es preciso
disipar esta ilusión, extendida por algunos pseudocientíficos. Sin duda se
podrían paliar ciertas taras genéticas, pero
solamente en el individuo que las padece,
no en su descendencia. No sólo la genética molecular moderna no nos propone ningún
medio de
actuar en el patrimonio hereditario para enriquecerle con nuevos rasgos, para
crear un «superhombre» genético, sino que revela la vanidad de una esperanza
así: la escala microscópica del genoma prohíbe por el momento, y sin duda para
siempre, tales manipulaciones. Aparte de las quimeras de ciencia ficción, el
único medio de «mejorar» la especie humana sería operar una selección deliberada
y severa. Pero, ¿quién querrá, quién osará emplearla?
El peligro, para la especie, de las condiciones de no selección, o de selección
al revés, que reinan en las sociedades avanzadas, es cierto. No llegará sin
embargo a ser realmente serio más que a largo plazo: digamos diez o quince
generaciones, varios siglos. Pero, las sociedades modernas están expuestas por
otro lado a amenazas más apremiantes y graves.
* * * *
No me refiero aquí a la explosión demográfica, a la destrucción de la
naturaleza, ni incluso a los megatones; sino a un mal mucho más profundo y
grave, a un mal del alma. Éste es el giro más grande de la evolución ideal que
la ha creado y sin cesar la agrava. El prodigioso desarrollo del conocimiento
desde hace tres siglos, constriñe hoy al hombre a una revisión desgarradora de
la concepción, arraigada desde hace decenas de miles de años, que él tenía de sí
mismo y de su relación con el universo.
Todo esto, sin embargo, el mal del alma y la potencia de los megatones, nos
viene de una idea simple: la naturaleza es objetiva, la verdad del conocimiento
no puede tener otra fuente que la confrontación sistemática de la lógica y de la
experiencia. No se acaba de comprender cómo no ha podido, en el reino de las
ideas, aquélla, tan simple y tan clara, aparecer con toda claridad más que cien
mil años después de la emergencia del Homo
sapiens;
cómo las más elevadas civilizaciones, la china por ejemplo, la ignoran, para
tomarla luego de Occidente; cómo, en el mismo Occidente, han sido precisos cerca
de 2.500 años, de Tales y Pitágoras a Galileo, Descartes, Bacon, para que, por
fin, ella se desprenda de la ganga que la encerraba en la pura práctica de las
artes mecánicas.
La selección de la ideas
Es tentador, para un biólogo, comparar la evolución de las ideas a la de la
biosfera. Porque el reino abstracto trasciende la biosfera más aún que ésta al
universo no vivo, las ideas han conservado algunas de las propiedades de los
organismos. Como éstos tienden a perpetuar su estructura y a multiplicarla,
pueden fusionar, recombinar, segregar su contenido y, en fin, evolucionar, y, en
esta evolución, la selección, sin ninguna duda, juega un gran papel. No me
aventuraría a proponer una teoría de la selección de las ideas. Pero se puede al
menos intentar definir algunos de los principales factores que desempeñan un
papel. Esta selección debe necesariamente operar a dos niveles: el del espíritu
y el de la performance.
El valor de performance de
una idea depende de la modificación de comportamiento que aporta al individuo o
al grupo que la adopta. Aquélla que confiera al grupo humano que la hace suya,
más cohesión, ambición, confianza en sí, le dará de hecho un aumento de poder de
expansión que asegurará la promoción de la misma idea. Este valor de promoción
no tiene necesariamente relación con la parte de verdad objetiva que la idea
pueda comportar. La poderosa armadura que para una sociedad constituye una
ideología religiosa no es debida a su estructura misma, sino al hecho de que
esta estructura es aceptada, que se impone. Por ello resulta muy difícil separar
el poder de invasión de una idea como ésta y su poder de performance.
El poder de invasión, en sí, es mucho más difícil de analizar. Digamos que
depende de las estructuras preexistentes del espíritu, entre las que se hallan
las ideas ya encaminadas por la cultura pero también, sin ninguna duda, ciertas
estructuras innatas que nos son, por otra parte, muy difíciles de identificar.
Pero se ve claramente que las ideas dotadas del más alto poder de invasión son
las que explican el
hombre asignándole su lugar en un destino inmanente, en cuyo seno se disuelve su
angustia.
La exigencia de explicación
Durante centenares de miles de años el destino de un hombre se confundía con el
de su grupo, de su tribu, fuera de la cual no podía sobrevivir. La tribu, en sí,
no podía sobrevivir ni defenderse más que por su cohesión. De ahí, el extremo
poder subjetivo de las leyes que organizaban y garantizaban esta cohesión. Algún
hombre podía a veces infringirlas; ninguno sin duda habría soñado con negarlas.
Viendo así la inmensa importancia selectiva que necesariamente asumieron tales
estructuras sociales, y durante tan largo periodo de tiempo, es difícil no
pensar que ellas debieron influenciar la evolución genética de las categorías
innatas del cerebro humano. Esta evolución debía no sólo facilitar la aceptación
de la ley tribal, sino crear la necesidad de
la explicación mítica que la cimenta, confiriéndole la soberanía. Nosotros somos
los descendientes de esos hombres. Es de ellos sin duda de quienes hemos
heredado la exigencia de una explicación, la angustia que nos constriñe a buscar
el sentido de la existencia. Angustia creadora de todos los mitos, de todas las
religiones, de todas las filosofías y de la ciencia misma.
Que esta imperiosa necesidad sea innata, inscrita de algún modo en el lenguaje
del código genético, que se desarrolle espontáneamente, no lo dudo por mi parte.
Fuera de la especie humana, no se encuentran en el reino animal organizaciones
sociales tan altamente diferenciadas, a no ser en algunos insectos: hormigas,
termitas o abejas. En los insectos sociales la estabilidad de las instituciones
no debe prácticamente nada a una herencia cultural, sino todo a la transmisión
genética. El comportamiento social es enteramente innato, automático.
En el hombre, las instituciones sociales, puramente culturales, no podrán jamás
alcanzar una estabilidad así; además ¿quién la desearía? La invención de los
mitos y de las religiones, la construcción de vastos sistemas filosóficos, son
el precio que el hombre debe pagar para sobrevivir como animal social sin caer
en un puro automatismo. Pero la herencia puramente cultural no sería bastante
segura, bastante poderosa por sí sola, para mantener las estructuras sociales.
Faltaba a esta herencia un soporte genético que se convirtiera en el alimento
exigido por el espíritu. Si ello no es así, ¿cómo explicar la universalidad, en
nuestra especie, del fenómeno religioso en la base de la estructura social?
¿Cómo explicar además que en la inmensa diversidad de mitos, religiones o
ideologías filosóficas, se encuentre la misma «forma» esencial?
Las ontogenias míticas y metafísicas
Es fácil ver que las «explicaciones», destinadas a fundar la ley aplacando la
angustia, son en su totalidad «historias» o, más exactamente, ontogenias. Los
mitos primitivos se refieren casi todos a héroes más o menos divinos cuya gesta
explica los orígenes del grupo y funda su estructura social sobre tradiciones
intocables: no se rehace la historia. Las grandes religiones tienen la misma
configuración; basándose en la historia de la vida de un profeta inspirado que,
si no es él mismo el fundador de todas las cosas, le representa, habla por él y
cuenta la historia de los hombres así como su destino. De todas las grandes
religiones, la judeocristiana es sin duda la más «primitiva» por su estructura
historicista, directamente ligada a la gesta de una tribu beduina, antes de ser
enriquecida por un profeta divino. El budismo, al contrario, más altamente
diferenciado, está ligado únicamente, en su forma original, al karma, la ley
transcendente que rige el destino individual. Es una historia de almas, más que
de hombres.
De Platón a Hegel y Marx, los grandes sistemas filosóficos proponen en su
totalidad ontogenias a la vez explicativas y normativas. En Platón, es cierto,
la ontogenia está al revés. En la historia él no ve más que corrupción gradual
de las formas ideales y, en la República,
es una máquina del tiempo lo que él, en suma, quiere poner en marcha.
Para Marx, como para Hegel, la historia se desarrolla según un plan inmanente,
necesario y favorable. El inmenso poder sobre los espíritus de la ideología
marxista, no es debido solamente a su promesa de una liberación del hombre, sino
también, y sin duda ante todo, a su estructura ontogénica, a la explicación que
da, entera y detallada, de la historia pasada, presente y futura. Sin embargo,
limitado a la historia humana e incluso adornado de las certidumbres de la
«ciencia», el materialismo histórico quedaba incompleto. Era preciso añadirle el
materialismo dialéctico, que aporta la interpretación total que exige el
espíritu: la historia humana y la del cosmos están asociadas como obedeciendo a
las mismas leyes eternas.
La ruptura de la «antigua alianza» animista y el mal del alma
moderna
Si es cierto que la necesidad de una explicación entera es innata, que su
ausencia es causa de profunda angustia; si la única forma de explicación, que
sabe aplacar la angustia, es la de una historia total que revele la
significación del hombre asignándole en los planes de la naturaleza un lugar
necesario; si para parecer verdadera, significante, apaciguante, la
«explicación» debe fundarse en la larga tradición animista,[55] se
comprende entonces por qué han sido precisos tantos milenios para que aparezcan
en el reino de las ideas las del conocimiento objetivo como única fuente
de verdad auténtica.
Esta idea austera y fría, que no propone ninguna explicación, pero que impone un
ascético renunciamiento a cualquier otro sustento espiritual, no podía calmar la
angustia innata; al contrario, la exasperaba. Ella pretendía, de un trazo,
borrar una tradición cien veces milenaria, asimilada a la misma naturaleza
humana; denunciaba la antigua alianza animista del hombre con la naturaleza, no
dejando, en el lugar de este precioso nexo, más que una búsqueda ansiosa en un
universo helado de soledad. ¿Cómo una idea así, que parecía estar revestida de
una puritana arrogancia, podía ser aceptada? No lo ha sido; no lo es aún. Y si a
pesar de todo se ha impuesto, es en razón, únicamente, de su prodigioso poder
de performance.
En tres siglos, la ciencia, fundada por el postulado de objetividad, ha
conquistado su lugar en la sociedad: en la práctica, mas no en las almas. Las
sociedades modernas están construidas sobre la ciencia. Le deben su riqueza, su
poderío y la certeza de que riquezas y poderes, aun mucho mayores, serán mañana,
si él lo quiere, accesibles al hombre. Pero también, igual que una «elección»
inicial en la evolución biológica de una especie puede comprometer el porvenir
de toda su descendencia, igual la elección, inconsciente en el origen, de una práctica científica
ha lanzado la evolución de la cultura por un camino de sentido único; trayecto
que el progresismo cientista del siglo XIX veía desembocar infaliblemente en una
expansión prodigiosa de la humanidad, mientras que hoy vemos abrirse delante
nuestro un abismo de tinieblas.
Las sociedades modernas han aceptado las riquezas y los poderes que la ciencia
les descubría. Pero no han aceptado, apenas han entendido, el profundo mensaje
de la ciencia: la definición de una nueva y única fuente de verdad, la exigencia
de una revisión total de los fundamentos de la ética, de una radical ruptura con
la tradición animista, el abandono definitivo de la «antigua alianza», la
necesidad de forjar una nueva. Armadas de todos los poderes, disfrutando de
todas las riquezas que deben a la Ciencia, nuestras sociedades intentan aún
vivir y enseñar sistemas de valores ya arruinados, en su raíz, por esta misma
ciencia.
Ninguna sociedad, antes de la nuestra, ha conocido semejante desgarramiento. En
las culturas primitivas como en las clásicas, las fuentes del conocimiento y las
de los valores eran confundidas por la tradición animista. Por primera vez en la
historia, una civilización intenta edificarse permaneciendo desesperadamente
ligada, para justificar sus valores, a la tradición animista, totalmente
abandonada como fuente de conocimiento, de verdad.
Las sociedades «liberales» de Occidente enseñan aún, con desdén, como base de su
moral, una repugnante mezcla de religiosidad judeocristiana, de progresismo
cientista, de creencia en los derechos «naturales» del hombre y de pragmatismo
utilitarista. Las sociedades marxistas profesaron siempre la religión
materialista y dialéctica de la historia; cuadro moral más sólido en apariencia
que el de las sociedades liberales, pero más vulnerable quizás en razón de la
misma rigidez que hasta ahora constituía su fuerza. Sea el que sea, todos estos
sistemas enraizados en el animismo están fuera del conocimiento objetivo, fuera
de la verdad, extraños y en definitiva hostiles a
la ciencia, que quieren utilizar, mas no respetar y servir. El divorcio es tan
grande, la mentira tan flagrante, que asedia y desgarra la conciencia de todo
hombre provisto de alguna cultura, dotado de alguna inteligencia y habitado por
esta ansiedad moral que es la fuente de toda creación. Es decir de todos
aquellos, entre los hombres, que llevan o llevarán las responsabilidades de la
sociedad y de la cultura en su evolución.
El mal del alma moderna es esta mentira, en la raíz del ser moral y social. Es
este mal, más o menos confusamente diagnosticado, que provoca el sentimiento de
temor, si no de odio, en cualquier caso de alienación, que hoy experimentan
tantos hombres respecto a la cultura científica. Lo más a menudo es hacia los
subproductos tecnológicos de la ciencia a los que se expresa abiertamente la
aversión: la bomba, la destrucción de la Naturaleza, la amenazadora demografía.
Es fácil, desde luego, replicar que la tecnología no es la ciencia y que, además
el empleo de la energía atómica será, pronto, indispensable para la
sobrevivencia de la humanidad; que la destrucción de la naturaleza denuncia una
tecnología insuficiente y no precisamente demasiada tecnología; que la explosión
demográfica es debida a que millones de niños son salvados de la muerte cada
año: ¿es preciso de nuevo dejarlos morir?
Discurso superficial, que confunde los signos con las causas profundas del mal.
Es claramente al mensaje esencial de la ciencia al que se dirige la repulsa. El
miedo está en el sacrilegio: en el atentado a los valores. Miedo enteramente
justificado. Es muy cierto que la ciencia atenta contra los valores. No
directamente, ya que no es juez y debe ignorarlos;
pero ella arruina todas las ontogenias míticas o filosóficas sobre las que la
tradición animista, de los aborígenes australianos a los dialécticos
materialistas, hace reposar los valores, la moral, los deberes, los derechos,
las prohibiciones.
Si acepta este mensaje en su entera significación, le es muy necesario al hombre
despertar de su sueño milenario para descubrir su soledad total, su radical
foraneidad. Él sabe ahora que, como un zíngaro, está al margen del universo
donde debe vivir. Universo sordo a su música, indiferente a sus esperanzas, a
sus sufrimientos y a sus crímenes.
Pero entonces ¿quién define el crimen? ¿Quién el bien y el mal? Todos los
sistemas tradicionales colocan la ética y los valores fuera del alcance del
hombre. Los valores no le pertenecen: ellos se imponen y es él quien les
pertenece. Él sabe ahora que ellos son sólo suyos y, al ser en fin el dueño, le
parece que se disuelven en el vacío indiferente del universo. Es entonces cuando
el hombre moderno se vuelve hacia, o mejor, contra la ciencia de la que calibra
ahora el terrible poder de destrucción, no sólo de los cuerpos, sino de la misma
alma.
Los valores y el conocimiento
¿Dónde está la solución? ¿Es preciso admitir definitivamente que la verdad
objetiva y la teoría de los valores constituyen para siempre terrenos opuestos,
impenetrables uno por el otro? Es la actitud que parecen tomar una gran parte de
los pensadores modernos, sean escritores, filósofos, o incluso hombres de
ciencia. Yo la creo no sólo inaceptable para la inmensa mayoría de los hombres,
sino absolutamente errónea, y ello por dos razones esenciales:
-
en primer lugar, desde luego, porque los valores y el conocimiento están
siempre y necesariamente asociados tanto en la acción como en el discurso;
-
a continuación y principalmente, porque la definición misma del
conocimiento «verdadero» se basa, en último término, en un postulado de orden
ético.
Cada uno de estos dos puntos necesita un pequeño desarrollo. La ética y el
conocimiento están inevitablemente ligados en la acción y por ella. La acción
pone en juego, o en cuestión, a
la vez el
conocimiento y los valores. Toda acción significa una ética, escoge o rechaza
ciertos valores; constituye unos valores escogidos, o lo pretende. Pero, por
otra parte, un conocimiento es necesariamente supuesto en toda acción, mientras
que, en compensación, la acción es una de las dos fuentes necesarias del
conocimiento.
En un sistema animista, la interpretación de la ética y del conocimiento no
plantea ningún conflicto, ya que el animismo evita toda distinción radical entre
estas dos categorías: las considera como dos aspectos de una misma realidad. La
idea de una ética social fundada sobre los «derechos» supuestos «naturales» del
hombre expresa una actitud tal que se revela también, pero de modo mucho más
sistemático y afirmado, en las tentativas de explicitación de la moral implícita
del marxismo.
Desde el momento en que se propone el postulado de objetividad, como condición
necesaria de toda verdad en el conocimiento, una distinción radical,
indispensable en la búsqueda de la verdad, es establecida entre el dominio de la
ética y el del conocimiento. El conocimiento en sí mismo es exclusivo de todo
juicio de valor (en tanto que «de valor epistemológico») mientras que la ética,
por esencia no
objetiva,
está por siempre excluida del campo del conocimiento.
En definitiva es esta distinción radical, propuesta como un axioma, la que ha
creado a la ciencia. Estoy tentado aquí de hacer notar que, si este
acontecimiento único en la historia de la cultura se produjo en el Occidente
cristiano antes que en el seno de otra civilización, es quizá, por una parte,
gracias al hecho de que la Iglesia reconocía una distinción fundamental entre el
dominio de lo sagrado y el de lo profano. Esta distinción no permitió a la
ciencia solamente buscar sus vías (a condición de no usurpar el dominio de lo
sagrado), sino que preparó al espíritu para la distinción mucho más radical que
proponía el principio de objetividad. Los occidentales pueden sentir cierta pena
al ver que para algunas religiones no existe, no puede existir, ninguna
distinción entre lo sagrado y lo profano. Para el hinduismo todo pertenece al
dominio sagrado; la misma noción de «profano» es incomprensible.
Esto no era más que un paréntesis. Volvamos al asunto. El postulado de
objetividad, al denunciar la «antigua alianza», impide al mismo tiempo toda
confusión entre juicios de conocimiento y juicios de valor. Pero sucede que
estas dos categorías están inevitablemente asociadas en la acción, comprendido
el discurso. Para permanecer fieles al principio, juzgaremos pues que todo
discurso (o acción) no debe ser considerado como significante, como auténtico,
más que si (o en la medida en que) explícita y conserva la distinción de las dos
categorías que él asocia. La noción de autenticidad deviene, así definida, el
dominio común donde se reúnen la ética y el conocimiento; donde los valores y la
verdad, asociados pero no confundidos, revelan su entera significación al hombre
atento que experimenta la resonancia. Por el contrario, el discurso inauténtico,
en el que las dos categorías se amalgaman y confunden, no puede conducir más que
a los contrasentidos más perniciosos, a las mentiras más criminales, aunque sean
inconscientes.
Se ve perfectamente que es en el discurso «político» (siempre interpreto
«discurso» en el sentido cartesiano) donde esta peligrosa amalgama se practica
de forma más constante y sistemática. Y ello no sólo por los políticos de
vocación. Los mismos hombres de ciencia, fuera de su dominio, se revelan a
menudo peligrosamente incapaces de distinguir la categoría de valores de la del
conocimiento.
Pero esto era otro paréntesis. Volvamos a las fuentes del conocimiento. El
animismo, hemos dicho, no quiere ni puede, por otra parte, establecer una
discriminación absoluta entre proposiciones de conocimiento y juicios de valor;
ya que, si una intención, por cuidadosamente disfrazada que esté, se supone
presente en el Universo, ¿qué sentido tendría una distinción así? En un sistema
objetivo, al contrario, toda confusión entre conocimiento y valores está prohibida.
Más (y éste es el punto esencial, la articulación lógica que asocia, en la raíz,
conocimiento y valores) esta prohibición, este «primer mandamiento» que funda el
conocimiento objetivo, no es en sí mismo y no sabría ser objetivo: es una regla
moral, una disciplina.
El conocimiento verdadero ignora los valores, pero hace falta para fundamentarlo
un juicio, o más bien un axioma de
valor. Es evidente que el plantear el postulado de objetividad como condición
del conocimiento verdadero constituye
una elección ética y no un juicio de conocimiento, ya que, según el mismo
postulado, no podía haber conocimiento «verdadero» con anterioridad a esta
elección arbitraria.
El postulado de objetividad, para establecer la norma del
conocimiento, define un valor que
es el mismo conocimiento objetivo. Aceptar el postulado de objetividad, es pues
enunciar la proposición de base de una ética: la
ética del conocimiento.
La ética del conocimiento
En la ética del conocimiento, es
la elección ética de un valor primitivo la que funda el conocimiento.
Por ello difiere radicalmente de las éticas animistas que en su totalidad se
consideran fundadas sobre el «conocimiento» de leyes inmanentes, religiosas o
«naturales», que se impondrían al hombre. La ética del conocimiento no se impone
al hombre; es
él, al contrario, quien se la impone haciendo
de ella axiomáticamente la
condición de autenticidad de todo discurso o de toda acción. El Discurso
del Método propone
una epistemología normativa, pero es preciso leerlo también y, ante todo, como
meditación moral, como ascesis del espíritu.
El discurso auténtico funda a su vez la ciencia y entrega a las manos de los
hombres los inmensos poderes que, hoy, le enriquecen y le amenazan, le liberan,
pero podrían también esclavizarle. Las sociedades modernas, tejidas por la
ciencia, viven de sus productos, han devenido dependientes como un toxicómano de
su droga. Ellas deben su poderío material a esta ética fundadora del
conocimiento, y su debilidad moral a los sistemas de valores, arruinados por el
mismo conocimiento, a los que intentan aún atenerse. Esta contradicción es
mortal. Es ella la que excava el abismo que vemos abrirse a nuestro paso. La
ética del conocimiento, creadora del mundo moderno, es la única compatible con
él, la única capaz, una vez comprendida y aceptada, de guiar su evolución.
* * * *
Comprendida y aceptada, ¿podrá serlo? Si es cierto, como creo, que la angustia
de la sociedad y la exigencia de una explicación total, apremiante, son innatas;
que esta herencia venida del fondo de las edades no es solamente cultural, sino
sin duda genética, ¿podemos pensar que esta ética austera, abstracta, orgullosa,
pueda calmar la angustia, saciar la exigencia? No lo sé. Mas quizá después de
todo no es totalmente imposible. ¿Puede ser que, más que una «explicación» que
la ética del conocimiento no sabría dar, el hombre necesite superación y
transcendencia? El poderío del gran sueño socialista, siempre vivo en las almas,
parece testimoniarlo a la perfección. Ningún sistema de valores puede pretender
constituir una verdadera ética, a menos de proponer un ideal que trascienda el
individuo al punto de justificar la necesidad por la que él se sacrifica.
Por la misma enjundia de su ambición, la ética del conocimiento podría quizá
satisfacer esta exigencia de superación. Ella define un valor transcendente, el
verdadero conocimiento, y propone al hombre no sólo servirse de él, sino en
adelante servirlo por una elección deliberada y consciente. Sin embargo, ella es
también un humanismo, ya que respeta en el hombre al creador y depositario de
esta trascendencia.
La ética del conocimiento es, igualmente, en un sentido, «conocimiento de la
ética», de los impulsos, de las pasiones, de las exigencias y de los límites del
ser biológico. En el hombre, ella sabe ver el animal, no sólo absurdo sino
extraño, precioso por su extrañeza misma, el ser que, perteneciendo
simultáneamente a dos reinos, la biosfera y el reino de las ideas, está a la vez
torturado y enriquecido por este dualismo desgarrador que se expresa tanto en el
arte y la poesía como en el amor humano.
La mayoría de los sistemas animistas, por el contrario, han querido ignorar,
envilecer o constreñir al hombre biológico, horrorizarle o aterrorizarle con
ciertos rasgos inherentes a su condición animal. La ética del conocimiento, por
el contrario, estimula al hombre a respetar y a asumir esta herencia, sabiendo,
cuando es necesario, dominarla. En cuanto a las más altas cualidades humanas, el
ánimo, el altruismo, la generosidad, la ambición creadora, la ética del
conocimiento, aun y reconociendo su origen socio-biológico, afirma también su
valor transcendente al servicio del ideal que ella define.
La ética del conocimiento y el ideal socialista
La ética del conocimiento, en fin, es, en mi opinión, la única actitud a la vez
racional y deliberadamente idealista sobre la que podría ser edificado un
verdadero socialismo. Este gran sueño del siglo XIX vive perennemente, en las
almas jóvenes, con una dolorosa intensidad. Dolorosa a causa de las traiciones
que este ideal ha sufrido y de los crímenes cometidos en su nombre. Es trágico,
pero quizás inevitable, que esta profunda aspiración no haya encontrado su
doctrina filosófica más que bajo la forma de una ideología animista. Es fácil
ver que el profetismo historicista fundamentado sobre el materialismo dialéctico
estaba, desde su nacimiento, cargado de todas las amenazas que han sido, en
efecto, realizadas. Más aún, quizá, que los demás animismos, el materialismo
histórico reposa sobre una confusión total de las categorías de valor y de
conocimiento. Es esta confusión la que le permite, con un discurso profundamente
inauténtico, proclamar que ha establecido «científicamente» las leyes de la
historia a las que el hombre no tiene otro recurso ni otro deber que obedecer,
si no quiere caer en la nada.
Una vez por todas, es preciso renunciar a esta ilusión que no es más que pueril
cuando no es mortal. ¿Cómo un socialismo auténtico podría construirse jamás
sobre una ideología inauténtica por esencia, burla de la ciencia sobre la que
pretende sinceramente, en el espíritu de sus adeptos, apoyarse? La sola
esperanza en el socialismo no está en una «revisión» de la ideología que lo
domina desde hace más de un siglo, sino en el abandono total de ella.
¿Dónde entonces encontrar la fuente de verdad y la inspiración moral de un
humanismo socialista realmente científico sino
en las fuentes de la misma ciencia, en la ética que funda el conocimiento,
haciendo de él, por libre elección, el valor supremo, medida y garantía de todos
los demás valores? Ética que funda la responsabilidad moral sobre la libertad de
esta elección axiomática. Aceptada como base de las instituciones sociales y
políticas, como medida de su autenticidad, de su valor, únicamente la ética del
conocimiento podría conducir al socialismo. Ella impone instituciones
consagradas a la defensa, a la extensión, al enriquecimiento del reino
transcendente de las ideas, del conocimiento, de la creación. Reino que habita
el hombre y en donde, cada vez más liberado de los apremios materiales y de las
servidumbres mentirosas del animismo, podría al fin vivir auténticamente,
protegido por instituciones que, viendo en él a la vez al sujeto y al creador
del reino, deberían servirle en su esencia más única y más preciosa.
Esto es quizás una utopía. Pero no es un sueño incoherente. Es una idea que se
impone por la sola fuerza de su coherencia lógica. Es la conclusión a la que
lleva necesariamente la búsqueda de la autenticidad. La antigua alianza ya está
rota; el hombre sabe al fin que está solo en la inmensidad indiferente del
Universo de donde ha emergido por azar. Igual que su destino, su deber no está
escrito en ninguna parte. Puede escoger entre el reino y las tinieblas.
Apéndices
Contenido:
1. Estructura de las proteínas
2. Ácidos nucleico
3. El código genético
4. Sobre la significación del segundo principio de la termodinámica
1. Estructura de las proteínas
Las proteínas son macromoléculas constituidas por la polimerización lineal de
cuerpos llamados «aminoácidos». La estructura general de la cadena
«polipeptídica» que resulta de esta polimerización es la siguiente:
En esta representación, los círculos blancos y negros y los cuadrados blancos
corresponden a diversos grupos de átomos (
CH; style="color: rgb(0, 0, 0); font-family: Verdana, Geneva, sans-serif; font-size: 16px; font-style: normal; font-variant-ligatures: normal; font-variant-caps: normal; font-weight: 400; letter-spacing: normal; orphans: 2; text-align: justify; text-indent: 0px; text-transform: none; white-space: normal; widows: 2; word-spacing: 0px; -webkit-text-stroke-width: 0px; text-decoration-thickness: initial; text-decoration-style: initial; text-decoration-color: initial"> =
CO; style="color: rgb(0, 0, 0); font-family: Verdana, Geneva, sans-serif; font-size: 16px; font-style: normal; font-variant-ligatures: normal; font-variant-caps: normal; font-weight: 400; letter-spacing: normal; orphans: 2; text-align: justify; text-indent: 0px; text-transform: none; white-space: normal; widows: 2; word-spacing: 0px; -webkit-text-stroke-width: 0px; text-decoration-thickness: initial; text-decoration-style: initial; text-decoration-color: initial"> =
NH), mientras que las letras R1,
R2,
etc., representan diferentes radicales orgánicos. Los 20 radicales de
aminoácidos que son constituyentes universales de las proteínas están
representados en la tabla 1.
Se ve que la cadena comprende tres tipos de unión entre átomos, o grupos de
átomos, a saber:
1º Entre círculo blanco y círculo negro (CH — CO).
2º Entre círculo blanco y cuadrado blanco (CH — NH).
3º Entre círculo negro y cuadrado blanco (CO — NH)
Tabla 1
Radicales aminoácidos
I) Hidrófobos
II) Hidrófilos
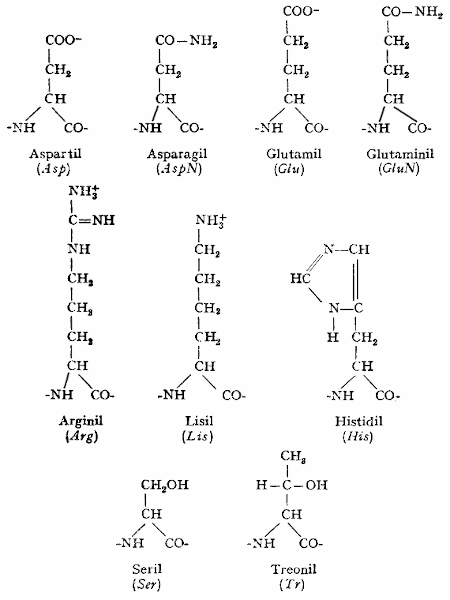
Esta última unión (llamada «peptídica») es rígida (trazo grueso en la figura
inicial de este apéndice): ella inmoviliza, uno respecto al otro, los átomos que
asocia. Al contrario, las otras dos uniones permiten una libre rotación (flechas
punteadas) de los átomos, uno respecto al otro. Esto permite a la fibra
polipeptídica plegarse sobre sí misma de modo extremadamente complejo y variado.
Sólo el impedimento de los átomos (principalmente los que constituyen los
radicales R1,
R2,
etc.) limita, en principio, estas posibilidades de plegamiento.
Sin embargo (ver cap. 5, Estructura
primaria y estructura globular de las proteínas),
en las proteínas globulares nativas, todas las moléculas de una misma especie
química (definida por la secuencia de
los radicales en la cadena) adoptan la misma configuración plegada. La figura 5
da esquemáticamente el recorrido de la cadena polipeptídica en una enzima, la
papaína. Se ve lo muy complejo que es este recorrido y, en apariencia, su
incoherencia.
Fig. 5. Representación esquemática de los pliegues de la cadena peptídica en
la molécula de papaína. J. Drenth, J. N. Jansonius, R. Koekoek, H. M. Swen y B.
G. Wolthers, «Nature», 218, págs. 929-932 (1968).
2. Ácidos nucleicos
Los ácidos nucleicos son macromoléculas que resultan de la polimerización lineal
de cuerpos llamados «nucleótidos». Éstos están constituidos por la asociación de
un azúcar con una base nitrogenada por una parte, y con un radical fosforilo por
otra. La polimerización tiene lugar por mediación de los grupos fosforilos que
asocian cada residuo de azúcar al precedente y al siguiente, formando así una
cadena «polinucleotídica».
En el ADN (ácido desoxirribonucleico) se encuentran cuatro nucleótidos que
difieren por la estructura de la base nitrogenada constituyente. Estas cuatro
bases, llamadas adenina, guanina, citosina y timina, se nombran en general A, G,
C y T. Son las letras del alfabeto genético. Por razones estéricas, la adenina
(A) en el ADN tiende a formar espontáneamente una asociación no covalente (ver
cap. 3, Uniones
covalentes y no covalentes)
con la timina (T) mientras que la guanina (G) se asocia con la citosina (C).
El ADN está constituido por dos fibras
polinucleotídicas asociadas por mediación de estas uniones no covalentes
específicas. En la doble fibra, A de una fibra está asociada a T de otra, G a C;
T a A y C a G. Las dos fibras son pues complementarias.
Esta estructura está esquemáticamente representada por la figura que viene a
continuación, donde los pentágonos simbolizan los radicales de azúcar, los
círculos negros los átomos de fósforo que aseguran la continuidad de cada una de
las dos cadenas, mientras que los cuadrados A, T, G, C representan las bases
emparejadas por pares (A-T; G-C; T-A; C-G) gracias a interacciones no
covalentes, indicadas en punteado. La estructura puede componerse de todas las
secuencias posibles de pares. No está limitada en cuanto a su longitud
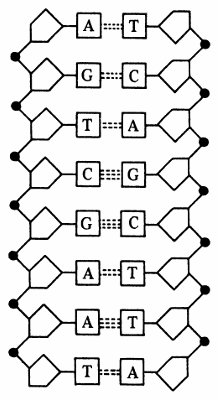
La replicación de
esta molécula procede por separación de las dos fibras, seguida por la
reconstitución, nucleótido a nucleótido, de las dos complementarias, lo que se
puede representar, con notación simplificada y limitándonos a cuatro pares, del
modo indicado en la siguiente figura.
Cada una de las dos moléculas así sintetizadas contiene una de las fibras de la
molécula madre y una fibra nueva formada por emparejamiento específico,
nucleótido. Estas dos moléculas son idénticas entre sí y también a la molécula
madre. Así es el mecanismo, muy simple en su principio, de la invariancia
replicativa.
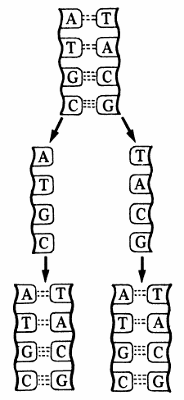
Las mutaciones resultan de los diversos tipos de accidentes que pueden afectar a
este mecanismo microscópico. Hoy en día se comprende bastante bien el mecanismo
químico de algunos de ellos. Por ejemplo, la sustitución de un par de
nucleótidos por otro es debida al hecho de que las bases nitrogenadas pueden,
además de su estado «normal», adoptar excepcional y transitoriamente una forma
tautomérica en la que la capacidad de emparejamiento específico de la base está
en cierto modo «invertido» (por ejemplo la base C, en la forma «excepcional», se
empareja con A y no con G). Se conocen agentes químicos que aumentan
considerablemente la probabilidad, es decir la frecuencia, de estos
emparejamientos «ilícitos». Estos agentes son poderosos «mutágenos».
Otros agentes químicos, capaces de insertarse entre los
nucleótidos en la fibra de ADN, la deforman y favorecen así los accidentes tales
como la eliminación o la adición de uno o varios nucleótidos.
En fin, las radiaciones ionizantes (rayos X y rayos cósmicos) provocan
especialmente diversos tipos de eliminaciones o de «mástiques».
3. El código genético
La estructura y las propiedades de una proteína están definidas por la secuencia
(el orden lineal) de los radicales aminoácidos en el polipéptido (cf. cap. 5, Formación
de las estructuras globulares).
Esta secuencia está determinada por la de los nucleótidos en un segmento de
fibra del ADN. El código genético (sensu
stricto)es
la regla que asocia, a una secuencia polinucleotídica dada, una secuencia
polipeptídica.
Como hay 20 residuos aminoácidos a especificar y sólo 4 «letras» (4 nucleótidos)
en el alfabeto del ADN, son precisos varios nucleótidos para especificar un
aminoácido. El código, de hecho, es a «tripletes»: cada aminoácido está
especificado por una secuencia de tres
nucleótidos.
Las correspondencias están dadas en la tabla 2.
Es preciso notar en primer lugar que la maquinaria de la traducción no utiliza
directamente las secuencias nucleotídicas del ADN, sino la «transcripción» de una de
las dos fibras a un polinucleótido, llamado «ácido ribonucleico mensajero» (ARN
mensajero). Los polinucleótidos del ARN difieren de los del ADN por algunos
detalles de estructura, principalmente por la sustitución de la base timina (T)
por la base uracilo (U).
Como es el ARN mensajero el que sirve de matriz para el ensamblaje secuencial de
los aminoácidos destinados a formar el polipéptido, el código figura en esta
tabla con su escritura en alfabeto ARN y no ADN.
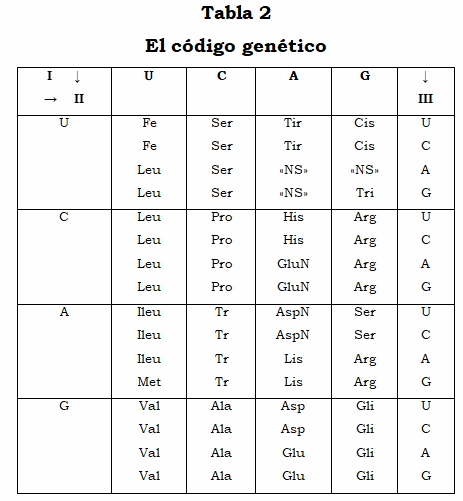
En esta tabla, la primera letra de cada triplete es leída en la columna vertical
de la izquierda, la segunda en la hilera horizontal, la tercera en la columna
vertical de la derecha. Los nombres de los radicales aminoácidos
correspondientes están resumidos (cd. Tabla de los radicales: Tabla 1, apéndice
1).
Se ve que, para la mayoría de los aminoácidos, existen varias notaciones
diferentes, bajo la forma de «tripletes» de nucleótidos. En un alfabeto de
cuatro letras se pueden en efecto formar 43 =
64 «palabras» de tres letras. Por ello no hay que especificar más que 20
residuos.
Por el contrario, tres tripletes (UAA, UAG, UGA) se nombran «NS» (non-sens =
sin sentido), porque no designan ningún aminoácido. Juegan sin embargo un papel
importante como signos de puntuación en la lectura de la secuencia nucleotídica.
El mecanismo de la traducción propiamente dicho es complejo; numerosos
constituyentes macromoleculares intervienen. El conocimiento de este mecanismo
no es indispensable para la inteligencia del texto. Basta con mencionar los
intermediarios que manejan en suma la clave de la traducción. Los ARN llamados
de «transferencia». Estas moléculas comportan en efecto:
-
Un grupo «aceptor» de aminoácidos; enzimas especiales reconocen por una
parte un aminoácido, por la otra un ARN de transferencia particular, y
catalizan la asociación (covalente) del aminoácido con la molécula de ARN.
-
Una secuencia complementaria de cada uno de los tripletes del
código, lo que permite a cada ARN de transferencia emparejarse al triplete
correspondiente del ARN mensajero.
Este emparejamiento tiene lugar en asociación con un constituyente complejo (el
ribosoma) que juega en suma el papel de «mesa de taller» donde se ensamblan los
diversos constituyentes del mecanismo. El ARN mensajero es leído secuencialmente,
mecanismo aún mal comprendido, que permite al ribosoma progresar, triplete a
triplete, a lo largo de la cadena polinucleotídica. Cada triplete a su vez se
empareja en la superficie del ribosoma con el ARN mensajero correspondiente,
portador del aminoácido especificado por este triplete. Una enzima cataliza en
cada etapa la formación de una unión peptídica entre el aminoácido que lleva el
ARN y el aminoácido precedente, en la extremidad de la cadena polipeptídica ya
formada, que aumenta así en una unidad. Después de ello, el ribosoma progresa un
triplete, y el proceso recomienza.
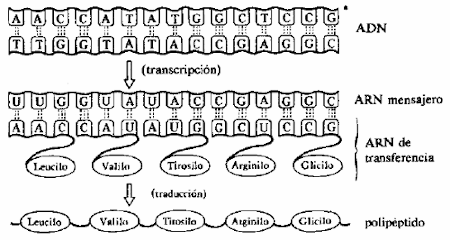
La figura anterior da un esquema de principio del mecanismo de transferencia de
la información correspondiente a una secuencia (arbitrariamente escogida) en el
ADN.
El ARN mensajero, en esta figura, se supone transcrito a partir de la fibra de
ADN marcada por un asterisco. En la realidad, los ARN de transferencia se
emparejan uno tras otro al mensajero. Para que resulte más claro, se los ha
colocado aquí emparejados simultáneamente.
4. Sobre la significación del segundo principio de la termodinámica
Se ha escrito tanto sobre la significación del segundo principio, sobre la
entropía, sobre la «equivalencia» entre la entropía negativa y la información,
que parece inútil volver a abordar este tema. Un repaso podrá sin embargo servir
a algunos lectores.
En su primera forma, puramente termodinámica (enunciada por Clausius en 1850,
como generalización del Teorema de Carnot), el segundo principio prevé que, en
un recinto energéticamente aislado,
todas las diferencias de temperatura deben tender a anularse espontáneamente.
O, y esto supone lo mismo, el principio estipula que, en el seno de un recinto
así, donde la temperatura sería uniforme, es imposible que aparezcan diferencias
de potencial térmico entre diferentes regiones del sistema. De donde proviene la
necesidad de gastar energía para enfriar una nevera, por ejemplo.
Entonces, en un recinto a temperatura uniforme, donde no subsiste ninguna
diferencia de potencial, no puede tener lugar ningún fenómeno (macroscópico). El
sistema es inerte.
Es en este sentido que se dice que el segundo principio prevé la degradación ineluctable
de la energía en el seno de un sistema aislado, como el Universo. La «entropía»
es la cantidad termodinámica que mide el
nivel de degradación de la energía de un sistema. Según el segundo principio,
por consecuencia, todo fenómeno, sea cual sea, es acompañado necesariamente de
un aumento de entropía en el seno del sistema donde acontece.
Es el desarrollo de la teoría cinética de la materia (o mecánica estática) el
que debía revelar la significación más profunda y más general del segundo
principio. La «degradación de la energía», o el aumento de entropía, es una
consecuencia estáticamente previsible de los movimientos y colisiones al azar de
las moléculas. Sean, por ejemplo, dos recintos a temperaturas diferentes,
puestos en comunicación uno con el otro. Las moléculas «calientes» (es decir
rápidas) y las moléculas «frías» (es decir lentas) van a pasar, en el azar de su
recorrido, de un recinto al otro, lo que anulará inevitablemente la diferencia
de temperatura entre los dos recintos. Se ve, gracias a este ejemplo, que el
aumento de entropía, en un sistema así, está ligado a un aumento de desorden: las
moléculas lentas y rápidas, primeramente separadas, están ahora mezcladas, y la
energía total del sistema se reparte estáticamente entre todas, debido a sus
colisiones; además, los dos recintos, al comienzo discernibles (por su
temperatura) se vuelven equivalentes. Antes de la mezcla, el sistema podía
realizar trabajo, ya que comportaba una diferencia de potencial entre los dos
recintos. Una vez alcanzado el equilibrio estático, ningún fenómeno puede
producirse ya en el seno del sistema.
Si el aumento de entropía mide el aumento del desorden en
un sistema, un aumento de orden corresponde a una disminución de entropía o,
como se prefiere a veces decir, a un enriquecimiento de entropía negativa. Sin
embargo, el grado de orden de un sistema es definible en otro lenguaje: el de la
información. El orden de un sistema, en este lenguaje, es igual a la cantidad de
información necesaria para la descripción de
este sistema. De donde proviene la idea, debida a Szilard y a Léon Brillouin, de
una cierta equivalencia entre «información» y «entropía negativa» (ver cap. 3, El
demonio de Maxwell).
Idea extremadamente fecunda, pero que puede dar lugar a generalizaciones o
asimilaciones imprudentes. Es sin embargo legítimo considerar que uno de los
enunciados fundamentales de la teoría de la información, a saber, que la
transmisión de un mensaje se acompaña necesariamente de una cierta disipación de
la información que contiene, es el equivalente, en informática, del segundo
principio en termodinámica.
Notas:
[1] En sentido propio, productos del arte, de la industria.
[2] En general, el autor, da a performance un
sentido próximo a logro, a ejecución conseguida. (N. del T.)
[3] Ver Apéndice, 4. Sobre la significación
del segundo principio de la termodinámica.
[4] Quizás haga falta subrayar que empleo aquí los
calificativos «animista» y «vitalista» con una acepción particular, algo
diferente del uso corriente.
[5] El pensamiento de Bergson no carece, desde luego, de
oscuridades y contradicciones aparentes. Parece que se pueda negar, por
ejemplo, que el dualismo bergsoniano sea esencial: ¿quizás haga falta
considerarlo como derivado de un monismo más primitivo? (C. Blanchard,
comunicación personal).
[6] Citemos también el siguiente texto de Henri Lefebvre (Le
matérialisme dialectique. PUF, París, 1949, p. 92): «La dialéctica,
lejos de ser un movimiento interior del espíritu, es real, antes que en el
espíritu, en el ser. Ella se impone al espíritu. Analicemos primeramente el
movimiento más simple y más abstracto: el del pensamiento más desnudo.
Descubriremos así las categorías más generales y su encadenamiento. Nos es
preciso luego ligar este movimiento al movimiento concreto, al contenido
dado: tomamos conciencia entonces del hecho de que el movimiento del
contenido y del ser se elucida para nosotros en las leyes dialécticas. Las
contradicciones en el pensamiento no vienen solamente del pensamiento, de su
impotencia o de su incoherencia definitivas, vienen también del contenido.
Su encadenamiento tiende hacia la expresión del movimiento
total del contenido y lo eleva al nivel de la conciencia y de la
reflexión.»
[7] «Llegamos pues a la conclusión de que, de una forma que
pertenecerá a los sabios del futuro darle luz, el calor irradiado en el
espacio debe necesariamente tener la posibilidad de convertirse en otra
forma de movimiento, bajo la cual pueda de nuevo concentrarse y volver a ser
activa. Así cae la dificultad esencial que se oponía a la reconversión de
soles muertos en nebulosas incandescentes. (…)
»Pero, sea cual sea la frecuencia y el inexorable rigor con los que este
ciclo se cumpla en el tiempo y en el espacio: sea cual sea el número de
millones de soles y de tierras que nazcan y perezcan; por mucho que sea el
tiempo que se necesite para que, en un sistema solar, las condiciones de
vida orgánica se establezcan, aunque sólo sea en un planeta; por
innumerables seres orgánicos que deban, en primer lugar, aparecer y perecer
antes que salga de su seno un animal con un cerebro capaz de pensar y que
encuentre por un corto lapso de tiempo las condiciones propias para la vida,
para ser luego exterminado también sin piedad; tenemos la certeza que, en
todas estas transformaciones, la materia permanece eternamente siendo la
misma, que ninguno de sus atributos puede jamás perderse y que, en
consecuencia, si ella debe en la Tierra exterminarse algún día, como
exigencia de una necesidad superior, su floración suprema, el espíritu
pensante, es preciso que por esa misma necesidad en otra parte y en otra
hora sea reproducido.» Engels, Dialéctica de la Naturaleza.
[8] Hay una simplificación deliberada. Ciertas estructuras
del ADN juegan un papel que es preciso considerar como teleonómico. Además
ciertos ARN (ácidos ribonucleicos) constituyen piezas esenciales de la
maquinaría que traduce del código genético (cf. Apéndice, 3. El
código genético). Sin embargo, proteínas específicas están igualmente
implicadas en estos mecanismos que, en casi todos los estados, ponen en
juego interacciones entre proteínas y ácidos nucleicos. La omisión de toda
discusión de estos mecanismos no afecta al análisis de las interacciones
teleonómicas moleculares y su interpretación general.
[9] Ver Apéndice, Tabla 1.
[10] Ver Apéndice, fig. 5.
[11] Los cuerpos que contienen un átomo de carbono unido a
cuatro grupos diferentes están por este hecho desprovistos de simetría. Se
les llama «ópticamente activos» porque al ser atravesados con la luz
polarizada se verifica una rotación en el plano de polarización, hacia la
izquierda (cuerpos levógiros: L) o hacia la derecha (cuerpos dextrógiros:
D).
[12] Recordemos que la energía de una unión es, por
definición, la que es preciso suministrar para romperla.
Pero, de hecho, la mayoría de las reacciones, principalmente bioquímicas,
consisten en el intercambio de uniones, más que su ruptura pura y
simple. La energía puesta en juego en una reacción es la que corresponde a
un intercambio del tipo:
AY + BX → AX + BY Ella es pues siempre inferior a la energía de ruptura.
[13] Se llama «metabolito» a todo cuerpo producido por el
metabolismo; «metabolitos esenciales» a los cuerpos universalmente
requeridos para el crecimiento y la multiplicación de las células.
[14] E. R. Stadtman, Advances in Enzymology,
28, 41-159 (1966). G. N. Cohen, Current Topics in Cellular Regulation,
I, 183-231 (1968).
[15] Se da el nombre de «ligando» a un cuerpo caracterizado
como tendente a ligarse a otro.
[16] J. Monod, J.-P. Changeux, y F. Jacob, «Journal of
Molecular Biology», 6, págs. 306-329 (1963).
[17] J. Monod, J. Wyman, y J.-P. Changeux, «Journal of
Molecular Biology», 12, págs. 88-118 (1965).
[18] Ver capitulo 3, Las proteínas-enzimas
como catalizadores específicos.
[19] El investigador finés Karstrom, que había en los años 30
aportado contribuciones notables al estudio de estos fenómenos, abandonó a
continuación la investigación, parece ser que para hacerse monje.
[20] F. Jacob y J. Monod, «Journal of Molecular Biology», 8,
págs. 318-356 (1961). Cf. igualmente «The lactose operon», Cold
Spring Harbour Monographv (1970), J. R. Beckwith y David Zipser, Edit.
[21] Cf. Beyond reductionism, Koestler y
Smythies, Ed. Hutchinson, Londres, 1969.
[22] Von Bertalanfy, en Koestler, loc. cit.
[23] Se sabe que la aparición de estructuras y de propiedades
nuevas en el curso del desarrollo embrionario ha sido a menudo calificada de
proceso «epigenético», como testimoniando un enriquecimiento gradual del
organismo a partir de la pura entrega genética, representada por el huevo
inicial. El adjetivo es a menudo empleado en referencia a teorías, hoy
superadas, que oponían los «preformacionistas» (que creían que el huevo
contenía una miniatura del animal adulto) a los epigenetistas (que creían en
un enriquecimiento real de la información inicial).
Empleo aquí este término para calificar, sin referencia a ninguna teoría,
todo proceso de desarrollo estructural y funcional.
[24] M. Nomura, «Ribosomes», «Scientific American», 221, 28
(1969).
[25] Se llaman «bacteriófagos» a los virus que atacan a las
bacterias.
[26] R. S. Edgar y W. B. Wood, «Morphogenesis of
bacteriophage T. in extracts of mutant infected cells», «Proceedings of the
National Academy of Science», 55, 498 (1966).
[27] J.-P. Changeux, en «Symmetry and function in biological
systems at the macromolecular level», A. Engström y B. Strandberg, edit., «Nobel
Symposium», N.º 11, págs. 235-256 (1969), John Wiley y Sons Inc., Nueva York.
[28] Cf. Popper: The Open Society and its
Enemies, Rutledge. Londres (1945).
[29] . Weisskopf, en Symmetry and Function in
biological Systems al the macromolecular level, Engström and Strandberg
Ed., «Nobel Symposium» N.º 11, págs. 28, Wiley y Sons, Nueva York (1969).
[30] Ver Apéndice, 2. Ácidos nucleicos.
[31] Replicación = Réplica, en el sentido de repetición, de
copia. Igual sentido para replicativo, replicado, etc. (N.
del T.)
[32] L. Orgel «Journal of Molecular Biology», 38, págs,
381-393 (1968).
[33] Se volverá sobre este punto en el capítulo 8.
[34] Explicación parcial, v. cap. 7, «Paradoja»
de la estabilidad de las especies.
[35] Orgel, L. E., Proceedings of the
National Academy of Science, 49, pág. 517 (1963).
[36] Cf. Apéndice, 2. Ácidos nucleicos.
[37] Simpson, The Meaning of Evolution.
Yale University Press (1967).
[38] Cf. N. Tinbergen, Social Behavior in
Animals, Methuen, Londres (1953)
[39] Leroi-Gourhan, Le geste et la parole,
Albin-Michel (1964). R. L. Holloway, «Current Anthropology», 10, 395 (1969)
- J. Bronowsky, en To Honor Roman Jakobson, Mouton, París, página 347
(1967).
[40] Según Leroi-Gourhan, loc. cit.
[41] B. T. Gardner y R. A. Gardner, en Behavior
of nonhuman Primates, Schrier y Stolnitz, Edit., Academic Press, Nueva
York (1970).
[42] E. Lenneberg, Biological Foundations of
Language, Wiley, Nueva York (1976).
[43] Lenneberg, loc. Cit.
[45] Cf. F. Crick «Journal of Molecular Biology», 38, págs.
367-279 (1968).
[47] Una teoría según la cual la memoria estaría codificada
en la secuencia de los radicales de algunas macromoléculas (ácidos
ribonucleicos) ha sido aceptada recientemente por algunos fisiológicos.
Éstos creen así aparentemente juntar y utilizar las concepciones sacadas del
estudio del código genético. Pero esta teoría es insostenible en relación,
precisamente, con nuestros conocimientos actuales sobre el código y los
mecanismos de la traducción.
[48] A excepción, quizá de las abejas.
[49] J. Z. Young, A Model of the Brain,
Oxford University Press (1964).
[50] H. B. Barlow, «Journal of Physiology», 119, págs. 69-88
(1953).
[51] T. N. Wiesel y D. H. Hubel «J. Neurophysiol.», 29,
páginas 1115-1156 (1966).
[52] D. H. Hubel y T. N. Wiesel, «Journal of Physiology»,
148, págs. 574-591 (1959).
[53] K. Lorenz, Evolution and Modification of
Behavior, University of Chicago Press, Chicago (1965).
[54] J. Levi-Agresti y R. W. Sperry, Proceedings
of the National Academy of Sciences, 61, pág. 1151 (1968).
[55] Quizá sea preciso subrayar de nuevo que empleo este
calificado en un sentido particular, definido en el cap. 2 (cf.El
progresismo cientista).
|